
Monosex Populations of the Giant Freshwater Prawn Macrobrachium rosenbergii—From a Pre-Molecular Start to the Next Generation Era


Abstract
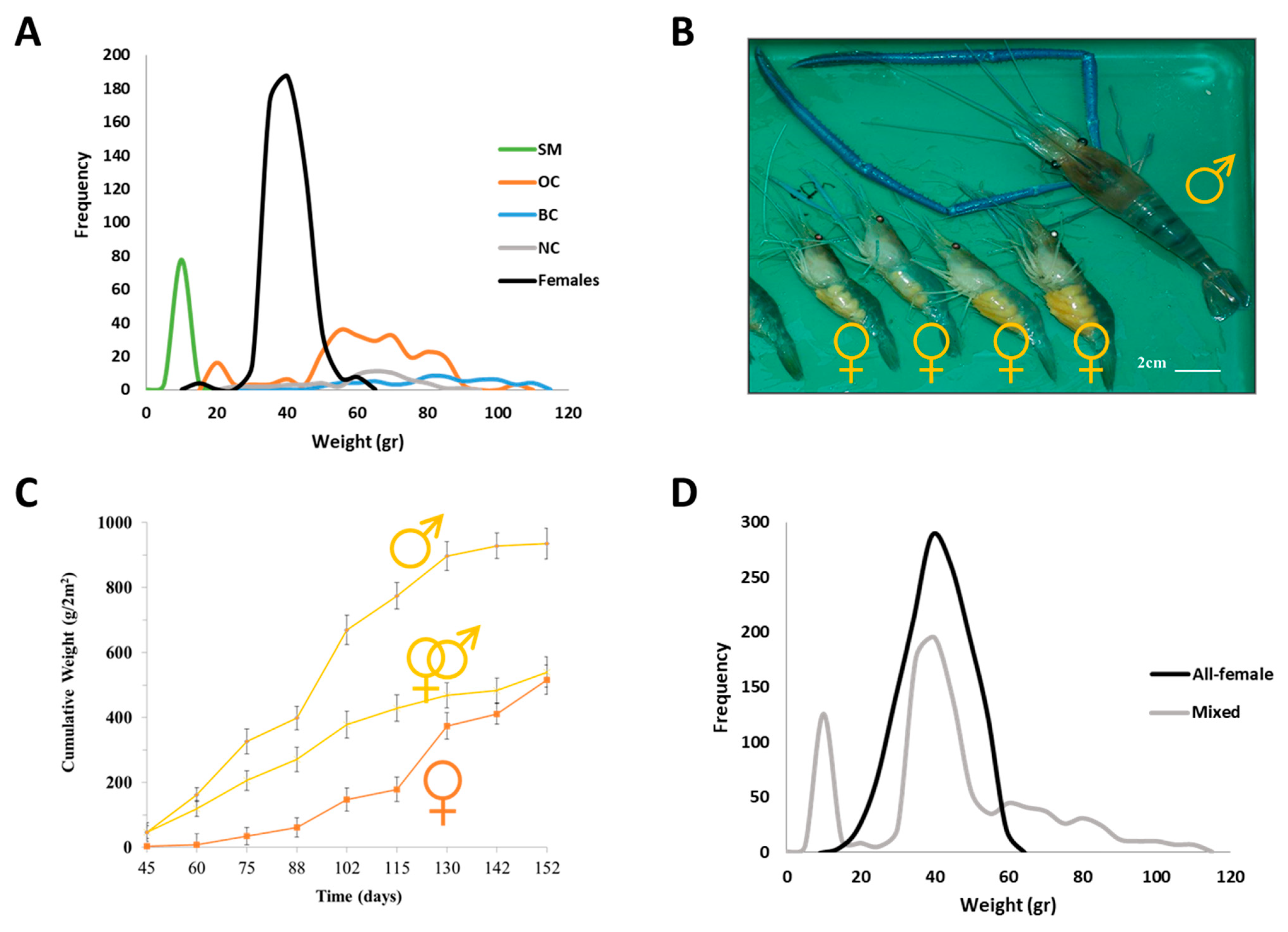
:1. The AG and IAG-Switch in Crustaceans
2. Manipulating the IAG-Switch
3. Controlling Elements of the IAG-Switch

{kind=link}
{kind=link}
{kind=link}
| Gene | Species | Tested Tissue or Stage | Sex Heritability Mechanism | Effect on Sexual Differentiation Processes | Reference |
|---|---|---|---|---|---|
| Dmrt11E | Macrobrachium rosenbergii | Post larvae | WZ/ZZ | MroDmrt11E knockdown induced a functional sex reversal | [85] |
| Terminal ampullae | WZ/ZZ | MroDmrt11E knockdown reduced MrIAG expression | [86] | ||
| Macrobrachium nipponense | Ovary and hepatopancreas (females) Abdominal ganglia (males) | WZ/ZZ | MniDMRT11E knockdown reduced MnVG expression in females and increased MnIAG expression in males | [87] | |
| Dsx | Fenneropenaeus chinensis | Male cephalothorax | WZ/ZZ | Two isoforms of FcIAG gene were down-regulated following FcDsx knockdown | [73] |
| Macrobrachium rosenbergii | AG | WZ/ZZ | MrDsx knockdown reduced MrIAG expression | [77] | |
| Cherax quadricarinatus | Cephalothoraxes of undifferentiated crayfish | WZ/ZZ | CqDsx knockdown decreased CqIAG expression | [76] | |
| SOXE | Portunus trituberculatus | AG and testis | XY/XX | PtSoxE siRNA reduced PtIAG and PtIR expression | [88] |
| SOXB2-1 | Eriocheir sinensis | Testis | WZ/ZZ | EsSoxB2-1 knockdown led to abnormal nucleus transformation during spermiogenesis | [89] |
| SOX9 | Scylla paramamosain | Cell culture | WZ/ZZ | Sox9 binding site mutation in SpVIH promoter reduced activity | [90] |
| CHH1 | Litopenaeus vannamei | AG | WZ/ZZ | Knockdown of either LvCHH1 or LvCHH2 resulted in increased LvIAG expression; injection of their recombinant protein led to decreased LvIAG expression. | [66] |
| CHH2 | |||||
| GC receptor | Litopenaeus vannamei | AG | WZ/ZZ | LvGC knockdown increased LvIAG expression | [66] |
| MIH | Macrobrachium nipponense | AG | WZ/ZZ | Gene knockdown increased IAG expression | [91] |
| GIH (VIH) | |||||
| Masc | Macrobrachium rosenbergii | Post larvae | WZ/ZZ | MrMasc knockdown obtained full sex reversal; Insulin-like signal pathway has been identified in MrMasc knockdown prawns. | [92] |
| Sxl | Cherax quadricarinatus | Undifferentiated juvenile prawns | WZ/ZZ | Sxl3 knockdown decreased CqIAG expression | [69] |
| Foxl2 | Scylla paramamosain | Ovaries | WZ/ZZ | Spfoxl2 knockdown increased SpVG expression | [93] |
| CFSH | Portunus trituberculatus | Eyestalk, AG and testis | XY/XX | PtCFSH knockdown increased PtIAG expression; injection of its recombinant protein led to decreased PtIAG expression. | [94] |
| Scylla paramamosain | Cultured AG | WZ/ZZ | Recombinant SpCFSH reduced SpIAG expression | [95] | |
| Cultured AG | WZ/ZZ | Recombinant SpCFSH reduced SpSTAT expression | [64] | ||
| STAT | Scylla paramamosain | Cultured AG | WZ/ZZ | SpSTAT knockdown reduced SpIAG expression | [64] |
| Tra2 | Macrobrachium nipponense | Gonads | WZ/ZZ | MnTra2 Inhibited MnSxl expression | [96] |
| GEM | Macrobrachium nipponense | AG and testis | WZ/ZZ | MnGEM knockdown increased MnIAG expression and testosterone content | [97] |
| BMP receptor | Scylla paramamosain | Pre/early/late vitellogenic females | WZ/ZZ | SpBMPR knockdown decreased vitellogenin receptor expression | [98] |
4. Distinct Biotechnologies for Monosex Populations of Prawns
5. Molecular and Genomic Implications
6. Future Environmental Applications of the IAG Switch
Supplementary Materials
Funding
Conflicts of Interest
References
- Wang, T.; He, K.; Blaney, L.; Chung, J.S. Testosterone and steroidogenic genes in the male blue crab Callinectes sapidus and their relationship with insulin-like androgenic gland factor (IAG) and crustacean female sex hormone (CFSH). Aquaculture 2023, 568, 739297. [Google Scholar] [CrossRef]
- Wang, T.; He, K.; Blaney, L.; Chung, J.S. 17β-Estradiol (E2) may be involved in the mode of crustacean female sex hormone (CFSH) action in the blue crab, Callinectes sapidus. Front. Endocrinol. 2022, 13, 962576. [Google Scholar] [CrossRef] [PubMed]
- Okumura, T.; Sakiyama, K. Hemolymph levels of vertebrate-type steroid hormones in female kuruma prawn Marsupenaeus japonicus (Crustacea: Decapoda: Penaeidae) during natural reproductive cycle and induced ovarian development by eyestalk ablation. Fish. Sci. 2004, 70, 372–380. [Google Scholar] [CrossRef]
- Lin, J.; Shi, X.; Fang, S.; Zhang, Y.; You, C.; Ma, H.; Lin, F. Comparative transcriptome analysis combining SMRT and NGS sequencing provides novel insights into sex differentiation and development in mud crab (Scylla paramamosain). Aquaculture 2019, 513, 734447. [Google Scholar] [CrossRef]
- Sugestya, I.N.G.; Widodo, M.S.; Soeprijanto, A. Effect of 17β-estradiol on feminization, growth rate and survival rate of pacific white shrimp (Litopenaeus vannamei, Boone 1931) postlarvae. J. Exp. Life Sci. 2018, 8, 37–42. [Google Scholar] [CrossRef]
- Yano, I. Effect of 17α-hydroxy-progesterone on vitellogenin secretion in kuruma prawn, Penaeus japonicus. Aquaculture 1987, 61, 49–57. [Google Scholar] [CrossRef]
- Yano, I. Induced ovarian maturation and spawning in greasyback shrimp, Metapenaeus ensis, by progesterone. Aquaculture 1985, 47, 223–229. [Google Scholar] [CrossRef]
- Liu, M.; Pan, J.; Liu, Z.; Cheng, Y.; Gong, J.; Wu, X. Effect of estradiol on vitellogenesis and oocyte development of female swimming crab, Portunus trituberculatus. Aquaculture 2018, 486, 240–245. [Google Scholar] [CrossRef]
- Cronin, L.E. Anatomy and histology of the male reproductive system of Callinectes sapidus Rathbun. J. Morphol. 1947, 81, 209–239. [Google Scholar] [CrossRef]
- Charniaux-Cotton, H. Découverte chez un Crustacé Amphipode (Orchestia gammarella) d’une glande endocrine responsable de la différenciation des caractères sexuels primaires et secondaires mâles. Comptes Rendus Hebd. Des Seances De L’academie Des Sci. Paris 1954, 239, 780–782. [Google Scholar]
- Charniaux-Cotton, H. Le determinisme hormonal des caracteres sexuels d’Orchestia gammarella (Crustace Amphipode). CR Acad. Sci. Paris 1955, 240, 1487–1489. [Google Scholar]
- Charniaux-Cotton, H. Croissance, régénération et déterminisme endocrinien des caractères sexuels d’Orchestia gammarella Pallas (Crustacé Amphipode). Ann. Sci. Nat. Zool. Biol. Anim. 1957, 19, 411–559. [Google Scholar]
- Nagamine, C.; Knight, A.W.; Maggenti, A.; Paxman, G. Masculinization of female Macrobrachium rosenbergii (de Man) (Decapoda, Palaemonidae) by androgenic gland implantation. Gen. Comp. Endocrinol. 1980, 41, 442–457. [Google Scholar] [CrossRef]
- Taketomi, Y.; Nishikawa, S. Implantation of androgenic glands into immature female crayfish, Procambarus clarkii, with masculinization of sexual characteristics. J. Crustac. Biol. 1996, 16, 232–239. [Google Scholar] [CrossRef]
- Cui, Z.; Liu, H.; Lo, T.S.; Chu, K.H. Inhibitory effects of the androgenic gland on ovarian development in the mud crab Scylla paramamosain. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2005, 140, 343–348. [Google Scholar] [CrossRef]
- Liu, H.; Cai, S.-L.; Zhang, C.-F.; Chu, K.-H. Masculinization of female Eriocheir sinensis by injecting the extract of androgenic gland of E. sinensis and Scylla paramamosain. J. Fish. China 2006, 30, 577–585. [Google Scholar]
- Katakura, Y. Partial purification and characterization of androgenic gland hormone from the isopod crustacean, Armadillidium vulgare. Annot. Zool. 1975, 48, 203–209. [Google Scholar]
- Katakura, Y.; Hasegawa, Y. Masculinization of females of the isopod crustacean, Armadillidium vulgare, following injections of an active extract of the androgenic gland. Gen. Comp. Endocrinol. 1983, 49, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, Y.; Haino-Fukushima, K.; Katakura, Y. Isolation and properties of androgenic gland hormone from the terrestrial isopod, Armadillidium vulgare. Gen. Comp. Endocrinol. 1987, 67, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Okuno, A.; Hasegawa, Y.; Nagasawa, H. Purification and properties of androgenic gland hormone from the terrestrial isopod Armadillidium vulgare. Zool. Sci. 1997, 14, 837–842. [Google Scholar] [CrossRef]
- Okuno, A.; Hasegawa, Y.; Ohira, T.; Katakura, Y.; Nagasawa, H. Characterization and cDNA cloning of androgenic gland hormone of the terrestrial isopod Armadillidium vulgare. Biochem. Biophys. Res. Commun. 1999, 264, 419–423. [Google Scholar] [CrossRef] [PubMed]
- Ohira, T.; Hasegawa, Y.; Tominaga, S.; Okuno, A.; Nagasawa, H. Molecular cloning and expression analysis of cDNAs encoding androgenic gland hormone precursors from two porcellionidae species, Porcellio scaber and P. dilatatus. Zool. Sci. 2003, 20, 75–81. [Google Scholar] [CrossRef]
- Godbillon, G.; Balesdent, M. Specific substance of steroid nature in the male genital tract of the decapod crustacean, Carcinus maenas. Comptes Rendus Seances L’academie Sci. 1974, 278, 2823–2826. [Google Scholar]
- Veith, W.; Malecha, S. Histochemical-study of the distribution of lipids, 3-alpha-hydroxysteroid and 3-beta-hydroxysteroid dehydrogenase in the androgenic gland of the cultured prawn, Macrobachium-rosenbergii (de man)(Crustacea, Decapoda). S. Afr. J. Sci. 1983, 79, 84–85. [Google Scholar]
- Manor, R.; Weil, S.; Oren, S.; Glazer, L.; Aflalo, E.D.; Ventura, T.; Chalifa-Caspi, V.; Lapidot, M.; Sagi, A. Insulin and gender: An insulin-like gene expressed exclusively in the androgenic gland of the male crayfish. Gen. Comp. Endocrinol. 2007, 150, 326–336. [Google Scholar] [CrossRef] [PubMed]
- Ventura, T.; Manor, R.; Aflalo, E.D.; Weil, S.; Raviv, S.; Glazer, L.; Sagi, A. Temporal silencing of an androgenic gland-specific insulin-like gene affecting phenotypical gender differences and spermatogenesis. Endocrinology 2009, 150, 1278–1286. [Google Scholar] [CrossRef] [PubMed]
- Levy, T.; Zupo, V.; Mutalipassi, M.; Somma, E.; Ruocco, N.; Costantini, M.; Abehsera, S.; Manor, R.; Chalifa-Caspi, V.; Sagi, A.; et al. Protandric Transcriptomes to Uncover Parts of the Crustacean Sex-Differentiation Puzzle. Front. Mar. Sci. 2021, 8, 745540. [Google Scholar] [CrossRef]
- Levy, T.; Tamone, S.L.; Manor, R.; Aflalo, E.D.; Sklarz, M.Y.; Chalifa-Caspi, V.; Sagi, A. The IAG-switch and further transcriptomic insights into sexual differentiation of a protandric shrimp. Front. Mar. Sci. 2020, 7, 587454. [Google Scholar] [CrossRef]
- Levy, T.; Rosen, O.; Simons, O.; Savaya Alkalay, A.; Sagi, A. The gene encoding the insulin-like androgenic gland hormone in an all-female parthenogenetic crayfish. PLoS ONE 2017, 12, e0189982. [Google Scholar] [CrossRef]
- Levy, T.; Sagi, A. The “IAG-switch”—A key controlling element in decapod crustacean sex differentiation. Front. Endocrinol. 2020, 11, 651. [Google Scholar] [CrossRef]
- Charniaux-Cotton, H.; Payen, G. Reproduction in malacostraca crustacea. In Endocrinology of Selected Invertebrate Types; Liss: New York, NY, USA, 1988; pp. 177–186. [Google Scholar]
- Ginsburger-Vogel, T.; Charniaux-Cotton, H. Sex determination. In The Biology of Crustacea; Bliss, D.E., Ed.; Academic Press: New York, NY, USA, 1982; pp. 257–281. [Google Scholar]
- Kuris, A.M.; Ra’anan, Z.; Sagi, A.; Cohen, D. Morphotypic differentiation of male Malaysian giant prawns, Macrobrachium rosenbergii. J. Crustac. Biol. 1987, 7, 219–237. [Google Scholar] [CrossRef]
- Sagi, A.; Ra’anan, Z.; Cohen, D.; Wax, Y. Production of Macrobrachium rosenbergii in monosex populations: Yield characteristics under intensive monoculture conditions in cages. Aquaculture 1986, 51, 265–275. [Google Scholar] [CrossRef]
- Sagi, A.; Cohen, D. Growth, maturation and progeny of sex-reversed Macrobrachium rosenbergii males. World Aquac. Rep. 1990, 21, 87–90. [Google Scholar]
- Nagamine, C.; Knight, A.W.; Maggenti, A.; Paxman, G. Effects of androgenic gland ablation on male primary and secondary sexual characteristics in the Malaysian prawn, Macrobrachium rosenbergii (de Man) (Decapoda, Palaemonidae), with first evidence of induced feminization in a nonhermaphroditic decapod. Gen. Comp. Endocrinol. 1980, 41, 423–441. [Google Scholar] [CrossRef] [PubMed]
- Aflalo, E.D.; Hoang, T.T.T.; Nguyen, V.; Lam, Q.; Nguyen, D.; Trinh, Q.; Raviv, S.; Sagi, A. A novel two-step procedure for mass production of all-male populations of the giant freshwater prawn Macrobrachium rosenbergii. Aquaculture 2006, 256, 468–478. [Google Scholar] [CrossRef]
- Agrawal, N.; Dasaradhi, P.; Mohmmed, A.; Malhotra, P.; Bhatnagar, R.K.; Mukherjee, S.K. RNA interference: Biology, mechanism, and applications. Microbiol. Mol. Biol. Rev. 2003, 67, 657–685. [Google Scholar] [CrossRef] [PubMed]
- Sagi, A.; Manor, R.; Ventura, T. Gene silencing in crustaceans: From basic research to biotechnologies. Genes 2013, 4, 620–645. [Google Scholar] [CrossRef]
- Lezer, Y.; Aflalo, E.D.; Manor, R.; Sharabi, O.; Abilevich, L.K.; Sagi, A. On the safety of RNAi usage in aquaculture: The case of all-male prawn stocks generated through manipulation of the insulin-like androgenic gland hormone. Aquaculture 2015, 435, 157–166. [Google Scholar] [CrossRef]
- Katakura, Y. Endocrine and genetic control of sex differentiation in the malacostracan Crustacea. Invertebr. Reprod. Dev. 1989, 16, 177–181. [Google Scholar] [CrossRef]
- Levy, T.; Rosen, O.; Eilam, B.; Azulay, D.; Aflalo, E.D.; Manor, R.; Shechter, A.; Sagi, A. A single injection of hypertrophied androgenic gland cells produces all-female aquaculture. Mar. Biotechnol. 2016, 18, 554–563. [Google Scholar] [CrossRef]
- Malecha, S.R.; Nevin, P.A.; Ha, P.; Barck, L.E.; Lamadrid-Rose, Y.; Masuno, S.; Hedgecock, D. Sex-ratios and sex-determination in progeny from crosses of surgically sex-reversed freshwater prawns, Macrobrachium rosenbergii. Aquaculture 1992, 105, 201–218. [Google Scholar] [CrossRef]
- Parnes, S.; Khalaila, I.; Hulata, G.; Sagi, A. Sex determination in crayfish: Are intersex Cherax quadricarinatus (Decapoda, Parastacidae) genetically females? Genet. Res. 2003, 82, 107–116. [Google Scholar] [CrossRef]
- Waiho, K.; Shi, X.; Fazhan, H.; Li, S.; Zhang, Y.; Zheng, H.; Liu, W.; Fang, S.; Ikhwanuddin, M.; Ma, H. High-density genetic linkage maps provide novel insights into ZW/ZZ sex determination system and growth performance in mud crab (Scylla paramamosain). Front. Genet. 2019, 10, 298. [Google Scholar] [CrossRef] [PubMed]
- Ventura, T.; Aflalo, E.; Weil, S.; Kashkush, K.; Sagi, A. Isolation and characterization of a female-specific DNA marker in the giant freshwater prawn Macrobrachium rosenbergii. Heredity 2011, 107, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Chandler, J.C.; Elizur, A.; Ventura, T. The decapod researcher’s guide to the galaxy of sex determination. Hydrobiologia 2018, 825, 61–80. [Google Scholar] [CrossRef]
- Ventura, T.; Manor, R.; Aflalo, E.D.; Weil, S.; Rosen, O.; Sagi, A. Timing sexual differentiation: Full functional sex reversal achieved through silencing of a single insulin-like gene in the prawn, Macrobrachium rosenbergii. Biol. Reprod. 2012, 86, 90. [Google Scholar] [CrossRef] [PubMed]
- Barki, A.; Karplus, I.; Khalaila, I.; Manor, R.; Sagi, A. Male-like behavioral patterns and physiological alterations induced by androgenic gland implantation in female crayfish. J. Exp. Biol. 2003, 206, 1791–1797. [Google Scholar] [CrossRef]
- Bardera, G.; Owen, M.A.; Façanha, F.N.; Sloman, K.A.; Alexander, M.E. The influence of sex on feeding behaviour in Pacific white shrimp (Litopenaeus vannamei). Appl. Anim. Behav. Sci. 2020, 224, 104946. [Google Scholar] [CrossRef]
- Nair, C.; Salin, K.; Raju, M.; Sebastian, M. Economic analysis of monosex culture of giant freshwater prawn (Macrobrachium rosenbergii De Man): A case study. Aquac. Res. 2006, 37, 949–954. [Google Scholar] [CrossRef]
- Phansawat, P.; Keetanon, A.; Rairat, T.; Pichitkul, P.; Poldetch, P.; Chuchird, N. Benefits of Male Monosex Culture of Giant Freshwater Prawn (Macrobrachium rosenbergii): Improving Growth Performance, Production Yield, and Profitability. J. Fish. Environ. 2022, 46, 157–168. [Google Scholar]
- Islam, S.S.; Golder, M.R.; Bir, J.; Mistry, S.K.; Siddique, M.N.A.; Ali, M.R.; Sabbir, W.; Azad, M.A.K.; Huq, K.A. Growth comparison of mono and mixed-sex giant freshwater prawn (Macrobrachium rosenbergii, De man) and stocking optimization of male monosex farming. Khulna Univ. Stud. 2022, 19, 43–54. [Google Scholar] [CrossRef]
- Malecha, S. The case for all-female freshwater prawn, Macrobrachium rosenbergii (De Man), culture. Aquac. Res. 2012, 43, 1038–1048. [Google Scholar] [CrossRef]
- Karplus, I.; Sagi, A. The biology and management of size variation. In Freshwater Prawns Biology and Farming; Wiley-Blackwell: Hoboken, NJ, USA, 2010; pp. 316–347. [Google Scholar]
- Rotem-Dai, N.; Weil, S.; Greenshpan, Y.; Abehsera, S.; Manor, R.; Ofir, R.; Gazit, R.; Rosental, B.; Aflalo, E.D.; Sagi, A. Lentiviral-Transduced Ectopic Expression of Androgenic Hormone in a Crustacean Hematopoietic Primary Cell Culture. Front. Mar. Sci. 2021, 8, 677679. [Google Scholar] [CrossRef]
- Rosen, O.; Manor, R.; Weil, S.; Gafni, O.; Linial, A.; Aflalo, E.D.; Ventura, T.; Sagi, A. A sexual shift induced by silencing of a single insulin-like gene in crayfish: Ovarian upregulation and testicular degeneration. PLoS ONE 2010, 5, e15281. [Google Scholar] [CrossRef] [PubMed]
- Ventura, T.; Manor, R.; Aflalo, E.D.; Weil, S.; Khalaila, I.; Rosen, O.; Sagi, A. Expression of an androgenic gland-specific insulin-like peptide during the course of prawn sexual and morphotypic differentiation. Int. Sch. Res. Not. Endocrinol. 2011, 2011, 476283. [Google Scholar] [CrossRef] [PubMed]
- Khalaila, I.; Manor, R.; Weil, S.; Granot, Y.; Keller, R.; Sagi, A. The eyestalk–androgenic gland–testis endocrine axis in the crayfish Cherax quadricarinatus. Gen. Comp. Endocrinol. 2002, 127, 147–156. [Google Scholar] [CrossRef]
- Sroyraya, M.; Chotwiwatthanakun, C.; Stewart, M.J.; Soonklang, N.; Kornthong, N.; Phoungpetchara, I.; Hanna, P.J.; Sobhon, P. Bilateral eyestalk ablation of the blue swimmer crab, Portunus pelagicus, produces hypertrophy of the androgenic gland and an increase of cells producing insulin-like androgenic gland hormone. Tissue Cell 2010, 42, 293–300. [Google Scholar] [CrossRef]
- Chung, J.S.; Manor, R.; Sagi, A. Cloning of an insulin-like androgenic gland factor (IAG) from the blue crab, Callinectes sapidus: Implications for eyestalk regulation of IAG expression. Gen. Comp. Endocrinol. 2011, 173, 4–10. [Google Scholar] [CrossRef]
- Keller, R. Crustacean neuropeptides: Structures, functions and comparative aspects. Experientia 1992, 48, 439–448. [Google Scholar] [CrossRef]
- Zhu, D.; Feng, T.; Mo, N.; Han, R.; Lu, W.; Shao, S.; Cui, Z. New insights for the regulatory feedback loop between type 1 crustacean female sex hormone (CFSH-1) and insulin-like androgenic gland hormone (IAG) in the Chinese mitten crab (Eriocheir sinensis). Front. Physiol. 2022, 13, 1054773. [Google Scholar] [CrossRef]
- Jiang, Q.; Lu, B.; Wang, G.; Ye, H. Transcriptional inhibition of Sp-IAG by crustacean female sex hormone in the mud crab, Scylla paramamosain. Int. J. Mol. Sci. 2020, 21, 5300. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Liu, A.; Zhu, Z.; Wang, Y.; Ye, H. Crustacean female sex hormone: More than a female phenotypes-related hormone in a protandric simultaneous hermaphroditism shrimp. Int. J. Biol. Macromol. 2023, 238, 124181. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Li, S.; Lv, X.; Xiang, J.; Manor, R.; Sagi, A.; Li, F. Sex-biased CHHs and their putative receptor regulate the expression of IAG gene in the shrimp Litopenaeus vannamei. Front. Physiol. 2019, 10, 1525. [Google Scholar] [CrossRef]
- Toyota, K.; Miyakawa, H.; Hiruta, C.; Sato, T.; Katayama, H.; Ohira, T.; Iguchi, T. Sex determination and differentiation in decapod and cladoceran crustaceans: An overview of endocrine regulation. Genes 2021, 12, 305. [Google Scholar] [CrossRef] [PubMed]
- Li, J. A review of sexual determination and differentiation in crustacean. J. Biosci. Med. 2022, 10, 19–37. [Google Scholar] [CrossRef]
- Zheng, J.; Cheng, S.; Jia, Y.; Gu, Z.; Li, F.; Chi, M.; Liu, S.; Jiang, W. Molecular identification and expression profiles of four splice variants of Sex-lethal gene in Cherax quadricarinatus. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2019, 234, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.; Wan, H.; Zhang, Z.; Sheng, Y.; Jia, X.; Wang, Y. Transcriptional regulation of IAG by dsx and foxl-2 in mud crab (Scylla paramamosain). Gen. Comp. Endocrinol. 2024, 345, 114396. [Google Scholar] [CrossRef] [PubMed]
- Peng, B.; Wan, H.; Zhang, Z.; Jia, X.; Liu, C.; Wang, Y. A novel Dmrt gene of crustacean: Functional analysis of idmrt-2 gene in the male reproductive system from Scylla paramamosain. Gene 2023, 850, 146922. [Google Scholar] [CrossRef]
- Wei, W.-Y.; Huang, J.-H.; Zhou, F.-L.; Yang, Q.-B.; Li, Y.-D.; Jiang, S.; Jiang, S.-G.; Yang, L.-S. Identification and Expression Analysis of Dsx and Its Positive Transcriptional Regulation of IAG in Black Tiger Shrimp (Penaeus monodon). Int. J. Mol. Sci. 2022, 23, 12701. [Google Scholar] [CrossRef]
- Li, S.; Li, F.; Yu, K.; Xiang, J. Identification and characterization of a doublesex gene which regulates the expression of insulin-like androgenic gland hormone in Fenneropenaeus chinensis. Gene 2018, 649, 1–7. [Google Scholar] [CrossRef]
- Zhang, P.; Yang, Y.; Xu, Y.; Cui, Z. Analyses of the Dmrt family in a decapod crab, Eriocheir sinensis uncover new facets on the evolution of DM domain genes. Front. Physiol. 2023, 14, 1201846. [Google Scholar] [CrossRef]
- Wang, L.; Liu, Z.-Q.; Ma, K.-Y.; Tao, J.-R.; Fang, X.; Qiu, G.-F. A testis-specific gene doublesex is involved in spermatogenesis and early sex differentiation of the Chinese mitten crab Eriocheir sinensis. Aquaculture 2023, 569, 739401. [Google Scholar] [CrossRef]
- Zheng, J.; Cai, L.; Jia, Y.; Chi, M.; Cheng, S.; Liu, S.; Li, F.; Gu, Z. Identification and functional analysis of the doublesex gene in the redclaw crayfish, Cherax quadricarinatus. Gene Expr. Patterns 2020, 37, 119129. [Google Scholar] [CrossRef] [PubMed]
- Zhong, P.; Zhou, T.; Zhang, Y.; Chen, Y.; Yi, J.; Lin, W.; Guo, Z.; Xu, A.; Yang, S.; Chan, S. Potential involvement of a DMRT family member (Mr-Dsx) in the regulation of sexual differentiation and moulting in the giant river prawn Macrobrachium rosenbergii. Aquac. Res. 2019, 50, 3037–3049. [Google Scholar] [CrossRef]
- Lv, Q.; Li, S.; Miao, M.; Jin, S.; Li, F. IAG Regulates the Expression of Cytoskeletal Protein-Encoding Genes in Shrimp Testis. Genes 2023, 14, 564. [Google Scholar] [CrossRef]
- Sun, R.; Yi, S.; Shi, L.; Tan, K.; Shi, R.; Yang, S.; Li, Y. Analysis of mRNA and MicroRNA Expression Profiles of Nervous Tissues and Reproductive Tissues in Male Procambarus clarkii After Silencing IAG. Mar. Biotechnol. 2023, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.; Glendinning, S.; Ventura, T. Transcriptomic Changes Following Induced De-Masculinisation of Australian Red Claw Crayfish Cherax quadricarinatus. Int. J. Mol. Sci. 2023, 24, 3292. [Google Scholar] [CrossRef]
- Lin, D.; Wang, M.; Yu, F.; Shi, W.; Luo, F.; Wu, C.; Yang, J.; Ma, W. Identification and expression pattern of three sex-related genes in the shrimp Neocaridina denticulata sinensis (Decapoda, Caridea). Crustaceana 2022, 95, 723–746. [Google Scholar] [CrossRef]
- Chen, Y.-L.; Wang, Y.-M.; Xu, H.-J.; Li, J.-W.; Luo, J.-Y.; Wang, M.-R.; Ma, W.-M. The characterization and knockdown of a male gonad-specific insulin-like receptor gene in the white shrimp Penaeus vannamei. Aquac. Rep. 2022, 27, 101345. [Google Scholar] [CrossRef]
- Qian, H.; Ma, K.; Feng, J.; Guo, Z.; Gong, J.; Chen, H.; Bai, H.; Qiu, G. Transcriptome analysis of the post-larvae of giant freshwater prawn (Macrobrachium rosenbergii) after IAG gene knockdown with microRNA interference. Gen. Comp. Endocrinol. 2022, 325, 114054. [Google Scholar] [CrossRef]
- Cohen, S.; Hasan, M.; Frishman, N.; Khalaila, I. A crustacean vitellogenin-derived peptide as an oocyte-specific delivery vehicle for gene silencing. Front. Mar. Sci. 2023, 10, 1128524. [Google Scholar] [CrossRef]
- Xu, H.-J.; Chen, Y.-L.; Wang, Y.-M.; Luo, J.-Y.; Li, J.-W.; Shen, S.-Q.; Yang, J.-S.; Ma, W.-M. Full functional sex reversal achieved through silencing of MroDmrt11E gene in Macrobrachium rosenbergii: Production of all-male monosex freshwater prawn. Front. Endocrinol. 2022, 12, 772498. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.-Q.; Ma, W.-M.; Zeng, Q.-G.; Qian, Y.-Q.; Yang, J.-S.; Yang, W.-J. Molecular cloning and sexually dimorphic expression of two Dmrt genes in the giant freshwater prawn, Macrobrachium rosenbergii. Agric. Res. 2014, 3, 181–191. [Google Scholar] [CrossRef]
- Wang, Y.; Jin, S.; Fu, H.; Qiao, H.; Sun, S.; Zhang, W.; Jiang, S.; Gong, Y.; Xiong, Y.; Wu, Y. Identification and characterization of the DMRT11E gene in the oriental river prawn Macrobrachium nipponense. Int. J. Mol. Sci. 2019, 20, 1734. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Xu, D.; Wang, M.; Xie, X.; Zhu, D. Molecular Characterization of a Male-specific SoxE Gene in the Swimming Crab, Portunus trituberculatus, and Transcriptional Interaction with Insulin-like Androgenic Gland Hormone. Fishes 2023, 8, 351. [Google Scholar] [CrossRef]
- Liu, Z.-Q.; Jiang, X.-H.; Qi, H.-Y.; Xiong, L.-W.; Qiu, G.-F. A novel SoxB2 gene is required for maturation of sperm nucleus during spermiogenesis in the Chinese mitten crab, Eriocheir sinensis. Sci. Rep. 2016, 6, 32139. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Jia, X.; Zou, Z.; Wang, X.; Wang, Y.; Zhang, Z. VIH from the mud crab is specifically expressed in the eyestalk and potentially regulated by transactivator of Sox9/Oct4/Oct1. Gen. Comp. Endocrinol. 2018, 255, 1–11. [Google Scholar] [CrossRef]
- Li, F.; Bai, H.; Zhang, W.; Fu, H.; Jiang, F.; Liang, G.; Jin, S.; Sun, S.; Qiao, H. Cloning of genomic sequences of three crustacean hyperglycemic hormone superfamily genes and elucidation of their roles of regulating insulin-like androgenic gland hormone gene. Gene 2015, 561, 68–75. [Google Scholar] [CrossRef]
- Luo, J.-Y.; Shen, S.-Q.; Xu, H.-J.; Yang, J.-S.; Ma, W.-M. The transcription factor masculinizer in sexual differentiation and achieved full functional sex reversal in prawn. iScience 2023, 26, 106968. [Google Scholar] [CrossRef]
- Wan, H.; Zhong, J.; Zhang, Z.; Xie, Y.; Wang, Y. Characterization of the foxl2 gene involved in the vtg expression in mud crab (Scylla paramamosain). Gene 2021, 798, 145807. [Google Scholar] [CrossRef]
- Wang, M.; Xu, R.; Tu, S.; Yu, Q.; Xie, X.; Zhu, D. Putative Role of CFSH in the Eyestalk-AG-Testicular Endocrine Axis of the Swimming Crab Portunus trituberculatus. Animals 2023, 13, 690. [Google Scholar] [CrossRef]
- Liu, A.; Liu, J.; Liu, F.; Huang, Y.; Wang, G.; Ye, H. Crustacean female sex hormone from the mud crab Scylla paramamosain is highly expressed in prepubertal males and inhibits the development of androgenic gland. Front. Physiol. 2018, 9, 924. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Fu, H.; Qiao, H.; Sun, S.; Zhang, W.; Jin, S.; Gong, Y.; Jiang, S.; Xiong, Y.; Wu, Y. RNA interference studies on the sex-linked genes transformer-2 and sex-lethal in the oriental river prawn Macrobrachium nipponense. Isr. J. Aquac. Bamidgeh 2017, 69. [Google Scholar] [CrossRef]
- Jin, S.; Hu, Y.; Fu, H.; Jiang, S.; Xiong, Y.; Qiao, H.; Zhang, W.; Gong, Y.; Wu, Y. Potential functions of Gem-associated protein 2-like isoform X1 in the oriental river prawn Macrobrachium nipponense: Cloning, qPCR, in situ hybridization, and RNAi analysis. Int. J. Mol. Sci. 2019, 20, 3995. [Google Scholar] [CrossRef] [PubMed]
- Shu, L.; Yang, Y.; Huang, H.; Ye, H. A bone morphogenetic protein ligand and receptors in mud crab: A potential role in the ovarian development. Mol. Cell. Endocrinol. 2016, 434, 99–107. [Google Scholar] [CrossRef]
- Levy, T.; Rosen, O.; Manor, R.; Dotan, S.; Azulay, D.; Abramov, A.; Sklarz, M.Y.; Chalifa-Caspi, V.; Baruch, K.; Shechter, A. Production of WW males lacking the masculine Z chromosome and mining the Macrobrachium rosenbergii genome for sex-chromosomes. Sci. Rep. 2019, 9, 12408. [Google Scholar] [CrossRef]
- Ventura, T.; Sagi, A. The insulin-like androgenic gland hormone in crustaceans: From a single gene silencing to a wide array of sexual manipulation-based biotechnologies. Biotechnol. Adv. 2012, 30, 1543–1550. [Google Scholar] [CrossRef]
- Shpak, N.; Manor, R.; Aflalo, E.D.; Sagi, A. Three generations of cultured prawn without W chromosome. Aquaculture 2017, 467, 41–48. [Google Scholar] [CrossRef]
- Levy, T.; Rosen, O.; Eilam, B.; Azulay, D.; Zohar, I.; Aflalo, E.D.; Benet, A.; Naor, A.; Shechter, A.; Sagi, A. All-female monosex culture in the freshwater prawn Macrobrachium rosenbergii–A comparative large-scale field study. Aquaculture 2017, 479, 857–862. [Google Scholar] [CrossRef]
- Molcho, J.; Levy, T.; Benet, A.; Naor, A.; Savaya, A.; Manor, R.; Abramov, A.; Aflalo, E.D.; Shechter, A.; Sagi, A. Three generations of prawns without the Z chromosome: Viable WW Macrobrachium rosenbergii all-female populations in polyculture with Oreochromis niloticus. Aquaculture 2020, 515, 734531. [Google Scholar] [CrossRef]
- Yuan, J.; Yu, Y.; Zhang, X.; Li, S.; Xiang, J.; Li, F. Recent advances in crustacean genomics and their potential application in aquaculture. Rev. Aquac. 2023, 15, 1501–1521. [Google Scholar] [CrossRef]
- González-Castellano, I.; Manfrin, C.; Pallavicini, A.; Martínez-Lage, A. De novo gonad transcriptome analysis of the common littoral shrimp Palaemon serratus: Novel insights into sex-related genes. BMC Genom. 2019, 20, 757. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Lv, J.; Lan, W.; Gao, B.; Liu, P. Discovery of Sex-Determining Genes in Portunus trituberculatus: A Comparison of Male and Female Transcriptomes During Early Developmental Stages. Front. Mar. Sci. 2022, 8, 811052. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, Y.; Li, S.; Zhang, X.; Xiang, J.; Li, F. Sex-Specific transcriptome sequencing of zoea I larvae and identification of sex-linked genes using bulked segregant analysis in Pacific white shrimp Litopenaeus vannamei. Mar. Biotechnol. 2020, 22, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Ventura, T.; Chandler, J.C.; Nguyen, T.V.; Hyde, C.J.; Elizur, A.; Fitzgibbon, Q.P.; Smith, G.G. Multi-Tissue transcriptome analysis identifies key sexual development-related genes of the ornate spiny lobster (Panulirus ornatus). Genes 2020, 11, 1150. [Google Scholar] [CrossRef]
- Grinshpan, N.; Abayed, F.A.A.; Wahl, M.; Ner-Gaon, H.; Manor, R.; Sagi, A.; Shay, T. The transcriptional landscape of the giant freshwater prawn: Embryonic development and early sexual differentiation mechanisms. Front. Endocrinol. 2022, 13, 1059936. [Google Scholar] [CrossRef]
- Abayed, F.A.A.; Manor, R.; Aflalo, E.D.; Sagi, A. Screening for Dmrt genes from embryo to mature Macrobrachium rosenbergii prawns. Gen. Comp. Endocrinol. 2019, 282, 113205. [Google Scholar] [CrossRef]
- Sharabi, O.; Manor, R.; Weil, S.; Aflalo, E.; Lezer, Y.; Levy, T.; Aizen, J.; Ventura, T.; Mather, P.; Khalaila, I. Identification and characterization of an insulin-like receptor involved in crustacean reproduction. Endocrinology 2016, 157, 928–941. [Google Scholar] [CrossRef]
- Gutekunst, J.; Andriantsoa, R.; Falckenhayn, C.; Hanna, K.; Stein, W.; Rasamy, J.; Lyko, F. Clonal genome evolution and rapid invasive spread of the marbled crayfish. Nat. Ecol. Evol. 2018, 2, 567–573. [Google Scholar] [CrossRef]
- Tan, M.H.; Gan, H.M.; Lee, Y.P.; Grandjean, F.; Croft, L.J.; Austin, C.M. A giant genome for a giant crayfish (Cherax quadricarinatus) with insights into cox1 pseudogenes in decapod genomes. Front. Genet. 2020, 11, 201. [Google Scholar] [CrossRef]
- Liu, B.-H.; Zhang, Y.-Q.; Ma, K.-Y.; Wu, X.-G.; Qiu, G.-F. Identification of sex-specific markers and ZW-chromosome DNA clones from the genomic BAC library of the Chinese mitten crab Eriocheir sinensis. Aquaculture 2022, 560, 738576. [Google Scholar] [CrossRef]
- Song, L.; Bian, C.; Luo, Y.; Wang, L.; You, X.; Li, J.; Qiu, Y.; Ma, X.; Zhu, Z.; Ma, L. Draft genome of the Chinese mitten crab, Eriocheir sinensis. GigaScience 2016, 5, s13742-13016-10112-y. [Google Scholar] [CrossRef] [PubMed]
- Veldsman, W.P.; Ma, K.Y.; Hui, J.H.L.; Chan, T.F.; Baeza, J.A.; Qin, J.; Chu, K.H. Comparative genomics of the coconut crab and other decapod crustaceans: Exploring the molecular basis of terrestrial adaptation. BMC Genom. 2021, 22, 313. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lv, J.; Liu, P.; Chen, P.; Wang, J.; Li, J. Genome survey and high-resolution backcross genetic linkage map construction of the ridgetail white prawn Exopalaemon carinicauda applications to QTL mapping of growth traits. BMC Genom. 2019, 20, 598. [Google Scholar] [CrossRef] [PubMed]
- Kenny, N.J.; Sin, Y.W.; Shen, X.; Zhe, Q.; Wang, W.; Chan, T.F.; Tobe, S.S.; Shimeld, S.M.; Chu, K.H.; Hui, J.H. Genomic sequence and experimental tractability of a new decapod shrimp model, Neocaridina denticulata. Mar. Drugs 2014, 12, 1419–1437. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yuan, J.; Sun, Y.; Li, S.; Gao, Y.; Yu, Y.; Liu, C.; Wang, Q.; Lv, X.; Zhang, X. Penaeid shrimp genome provides insights into benthic adaptation and frequent molting. Nat. Commun. 2019, 10, 356. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Zhang, X.; Liu, C.; Yu, Y.; Wei, J.; Li, F.; Xiang, J. Genomic resources and comparative analyses of two economical penaeid shrimp species, Marsupenaeus japonicus and Penaeus monodon. Mar. Genom. 2018, 39, 22–25. [Google Scholar] [CrossRef]
- Polinski, J.M.; Zimin, A.V.; Clark, K.F.; Kohn, A.B.; Sadowski, N.; Timp, W.; Ptitsyn, A.; Khanna, P.; Romanova, D.Y.; Williams, P. The American lobster genome reveals insights on longevity, neural, and immune adaptations. Sci. Adv. 2021, 7, eabe8290. [Google Scholar] [CrossRef]
- Anzalone, A.V.; Koblan, L.W.; Liu, D.R. Genome editing with CRISPR–Cas nucleases, base editors, transposases and prime editors. Nat. Biotechnol. 2020, 38, 824–844. [Google Scholar] [CrossRef]
- Gao, Y.; Zhang, X.; Zhang, X.; Yuan, J.; Xiang, J.; Li, F. CRISPR/Cas9-mediated mutation reveals Pax6 is essential for development of the compound eye in Decapoda Exopalaemon carinicauda. Dev. Biol. 2020, 465, 157–167. [Google Scholar] [CrossRef]
- Nakanishi, T.; Kato, Y.; Matsuura, T.; Watanabe, H. CRISPR/Cas-mediated targeted mutagenesis in Daphnia magna. PLoS ONE 2014, 9, e98363. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.; Serano, J.M.; Jarvis, E.; Bruce, H.S.; Wang, J.; Ray, S.; Barker, C.A.; O’Connell, L.C.; Patel, N.H. CRISPR/Cas9 mutagenesis reveals versatile roles of Hox genes in crustacean limb specification and evolution. Curr. Biol. 2016, 26, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Gui, T.; Zhang, J.; Song, F.; Sun, Y.; Xie, S.; Yu, K.; Xiang, J. CRISPR/Cas9-mediated genome editing and mutagenesis of EcChi4 in Exopalaemon carinicauda. G3 Genes Genomes Genet. 2016, 6, 3757–3764. [Google Scholar] [CrossRef]
- Molcho, J.; Manor, R.; Shamsian, M.; Sudarshan, G.; Ofir, R.; Parker, D.; Weil, S.; Wattad, H.; Hayun, E.; Levy, T. On genome editing in embryos and cells of the freshwater prawn Macrobrachium rosenbergii. Aquaculture 2022, 558, 738391. [Google Scholar] [CrossRef]
- Miao, M.; Li, S.; Yuan, J.; Liu, P.; Fang, X.; Zhang, C.; Zhang, X.; Li, F. CRISPR/Cas9-mediated gene mutation of EcIAG leads to sex reversal in the male ridgetail white prawn Exopalaemon carinicauda. Front. Endocrinol. 2023, 14, 1266641. [Google Scholar] [CrossRef]
- Stotish, R.L. Navigating the FDA approval process for genetically engineered animals. Transgenic Res 2014, 23, 198–199. [Google Scholar]
- Bratlie, S.; Halvorsen, K.; Myskja, B.K.; Mellegård, H.; Bjorvatn, C.; Frost, P.; Heiene, G.; Hofmann, B.; Holst-Jensen, A.; Holst-Larsen, T. A novel governance framework for GMO: A tiered, more flexible regulation for GMO s would help to stimulate innovation and public debate. EMBO Rep. 2019, 20, e47812. [Google Scholar] [CrossRef]
- Gherardi, F.; Aquiloni, L.; Diéguez-Uribeondo, J.; Tricarico, E. Managing invasive crayfish: Is there a hope? Aquat. Sci. 2011, 73, 185–200. [Google Scholar] [CrossRef]
- Kolar, C.S.; Lodge, D.M. Progress in invasion biology: Predicting invaders. Trends Ecol. Evol. 2001, 16, 199–204. [Google Scholar] [CrossRef]
- Gherardi, F. Crayfish invading Europe: The case study of Procambarus clarkii. Mar. Freshw. Behav. Physiol. 2006, 39, 175–191. [Google Scholar] [CrossRef]
- Savaya, A.; De Leo, G.; Aalto, E.; Levy, T.; Rosen, O.; Manor, R.; Aflalo, E.D.; Tricarico, E.; Sagi, A. The IAG gene in the invasive crayfish Procambarus clarkii–towards sex manipulations for biocontrol and aquaculture. Manag. Biol. Invasions 2020, 11, 237. [Google Scholar] [CrossRef]
- Erickson, R.A.; Thompson, H.M.; Kageyama, S.A.; Andriacchi, G.M.; Cupp, A.R.; Patiño, R.; Amberg, J.J. Assessing the suitability of YY males and ZZ females as an invasive species population control method across life histories. Biol. Invasions 2023, 25, 3737–3751. [Google Scholar] [CrossRef]
- Dearden, P.K.; Gemmell, N.J.; Mercier, O.R.; Lester, P.J.; Scott, M.J.; Newcomb, R.D.; Buckley, T.R.; Jacobs, J.M.; Goldson, S.G.; Penman, D.R. The potential for the use of gene drives for pest control in New Zealand: A perspective. J. R. Soc. N. Z. 2018, 48, 225–244. [Google Scholar] [CrossRef]
- Gierus, L.; Birand, A.; Bunting, M.D.; Godahewa, G.I.; Piltz, S.G.; Oh, K.P.; Piaggio, A.J.; Threadgill, D.W.; Godwin, J.; Edwards, O. Leveraging a natural murine meiotic drive to suppress invasive populations. Proc. Natl. Acad. Sci. USA 2022, 119, e2213308119. [Google Scholar] [CrossRef] [PubMed]
- Hammond, A.; Galizi, R.; Kyrou, K.; Simoni, A.; Siniscalchi, C.; Katsanos, D.; Gribble, M.; Baker, D.; Marois, E.; Russell, S. A CRISPR-Cas9 gene drive system targeting female reproduction in the malaria mosquito vector Anopheles gambiae. Nat. Biotechnol. 2016, 34, 78–83. [Google Scholar] [CrossRef]
- Sokolow, S.H.; Jones, I.J.; Jocque, M.; La, D.; Cords, O.; Knight, A.; Lund, A.; Wood, C.L.; Lafferty, K.D.; Hoover, C.M. Nearly 400 million people are at higher risk of schistosomiasis because dams block the migration of snail-eating river prawns. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160127. [Google Scholar] [CrossRef]
- Savaya-Alkalay, A.; Ndao, P.D.; Jouanard, N.; Diane, N.; Aflalo, E.D.; Barki, A.; Sagi, A. Exploitation of reproductive barriers between Macrobrachium species for responsible aquaculture and biocontrol of schistosomiasis in West Africa. Aquac. Environ. Interact. 2018, 10, 487–499. [Google Scholar] [CrossRef]
- Acosta-Pérez, V.J.; Ángeles-Hernández, J.C.; Vega-Sánchez, V.; Zepeda-Velázquez, A.P.; Añorve-Morga, J.; Ponce-Noguez, J.B.; Reyes-Rodríguez, N.E.; De-La-Rosa-Arana, J.L.; Ramírez-Paredes, J.G.; Gómez-De-Anda, F.R. Prevalence of Parasitic Infections with Zoonotic Potential in Tilapia: A Systematic Review and Meta-Analysis. Animals 2022, 12, 2800. [Google Scholar] [CrossRef]
- Lee, C.; Fielder, D. Maintenance and reproductive behaviour in the freshwatre prawn Macrobrachium australiense Holthuis (Crustacea: Decapoda: Palaemonidae). Mar. Freshw. Res. 1982, 33, 629–646. [Google Scholar] [CrossRef]
- Roberts, J.K.; Kuris, A. Predation and control of laboratory populations of the snail Biomphalaria glabrata by the freshwater prawn Macrobrachium rosenbergii. Ann. Trop. Med. Parasitol. 1990, 84, 401–412. [Google Scholar] [CrossRef]
- Savaya-Alkalay, A.; Ovadia, O.; Barki, A.; Sagi, A. Size-selective predation by all-male prawns: Implications for sustainable biocontrol of snail invasions. Biol. Invasions 2018, 20, 137–149. [Google Scholar] [CrossRef]
- Sokolow, S.H.; Lafferty, K.D.; Kuris, A.M. Regulation of laboratory populations of snails (Biomphalaria and Bulinus spp.) by river prawns, Macrobrachium spp.(Decapoda, Palaemonidae): Implications for control of schistosomiasis. Acta Trop. 2014, 132, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Savaya, A.; Glassner, H.; Livne-Luzon, S.; Chishinski, R.; Molcho, J.; Aflalo, E.D.; Zilberg, D.; Sagi, A. Prawn monosex populations as biocontrol agents for snail vectors of fish parasites. Aquaculture 2020, 520, 735016. [Google Scholar] [CrossRef]
- Azmi, W.A.; Khoo, S.C.; Ng, L.C.; Baharuddin, N.; Abd Aziz, A.; Ma, N.L. The current trend in biological control approaches in the mitigation of golden apple snail Pomacea spp. Biol. Control 2022, 175, 105060. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Lv, H.; Sheng, Q. Efficacy of Giant River Prawn Macrobrachium rosenbergii in Controlling the Invasive Snail Pomacea canaliculata: Implications for Ecological Farming. Diversity 2023, 15, 1001. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wahl, M.; Levy, T.; Ventura, T.; Sagi, A. Monosex Populations of the Giant Freshwater Prawn Macrobrachium rosenbergii—From a Pre-Molecular Start to the Next Generation Era. Int. J. Mol. Sci. 2023, 24, 17433. https://doi.org/10.3390/ijms242417433
Wahl M, Levy T, Ventura T, Sagi A. Monosex Populations of the Giant Freshwater Prawn Macrobrachium rosenbergii—From a Pre-Molecular Start to the Next Generation Era. International Journal of Molecular Sciences. 2023; 24(24):17433. https://doi.org/10.3390/ijms242417433
Chicago/Turabian StyleWahl, Melody, Tom Levy, Tomer Ventura, and Amir Sagi. 2023. "Monosex Populations of the Giant Freshwater Prawn Macrobrachium rosenbergii—From a Pre-Molecular Start to the Next Generation Era" International Journal of Molecular Sciences 24, no. 24: 17433. https://doi.org/10.3390/ijms242417433