
Evolution of the Critically Endangered Green Sawfish Pristis zijsron (Rhinopristiformes, Pristidae), Inferred from the Whole Mitochondrial Genome
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Sequence Analysis
2.2. Phylogenetic Analysis
2.3. Divergence Time Analysis
3. Results and Discussion
3.1. Mitochondrial Genome Structure and Organization
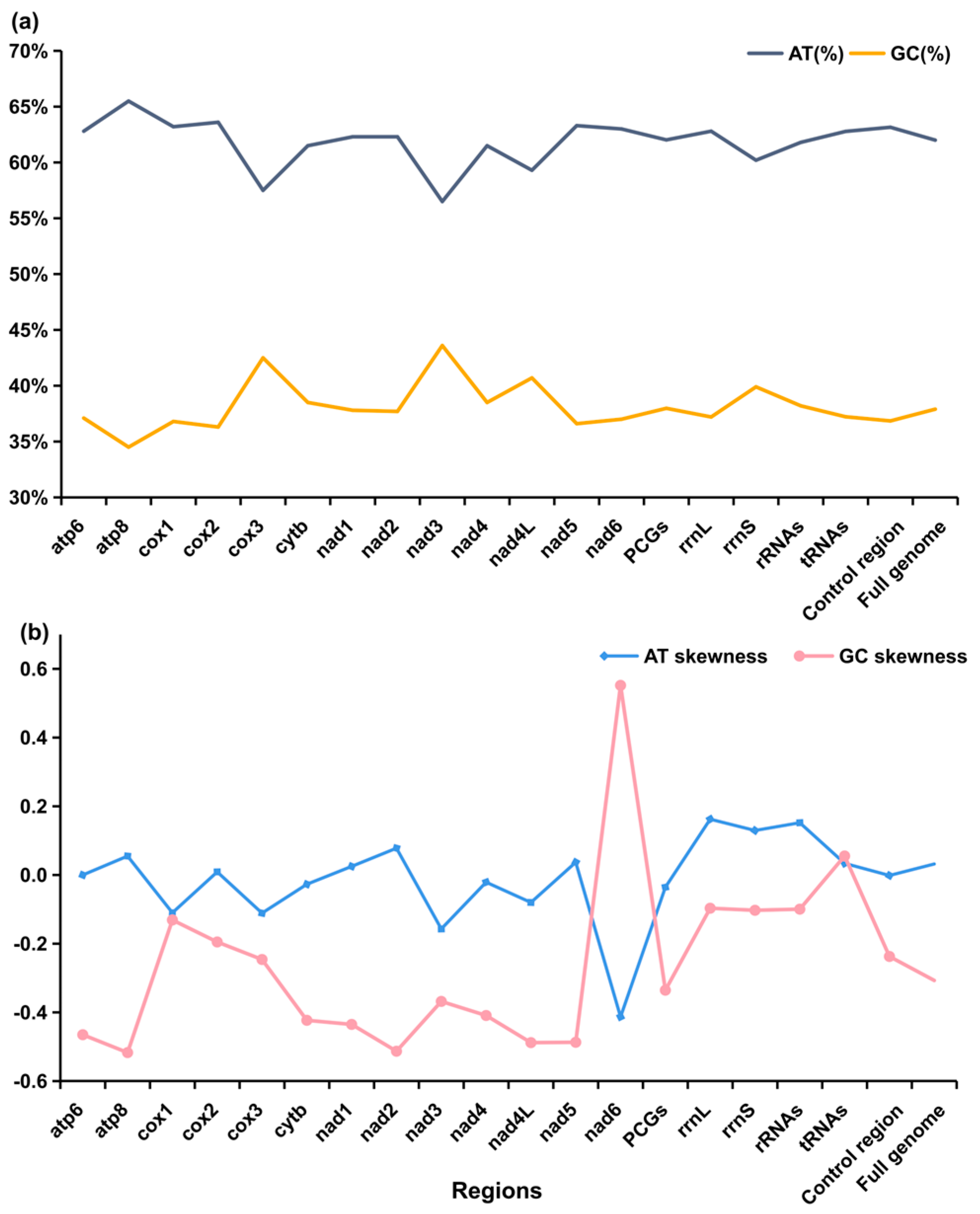
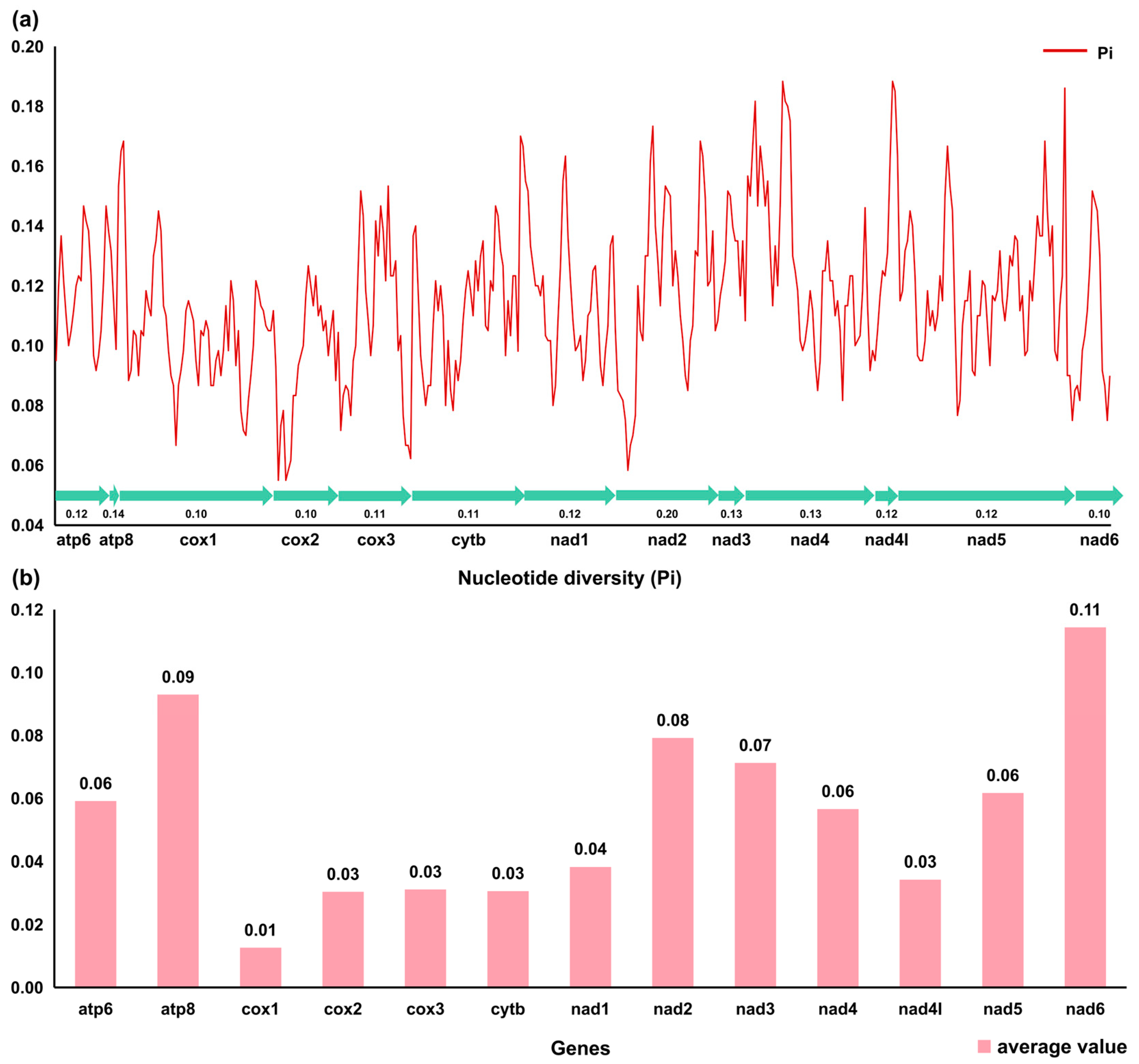
3.2. Protein Coding Genes and Codon Usage
3.3. Transfer RNAs, Ribosomal RNAs, and the D-Loop
3.4. Phylogenetic Inference
3.5. Divergence Time Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dulvy, N.K.; Pacoureau, N.; Rigby, C.L.; Pollom, R.A.; Jabado, R.W.; Ebert, D.A.; Finucci, B.; Pollock, C.M.; Cheok, J.; Derrick, D.H.; et al. Overfishing drives over one-third of all sharks and rays toward a global extinction crisis. Curr. Biol. 2021, 31, 4773–4787.e4778. [Google Scholar] [CrossRef] [PubMed]
- Stein, R.W.; Mull, C.G.; Kuhn, T.S.; Aschliman, N.C.; Davidson, L.N.K.; Joy, J.B.; Smith, G.J.; Dulvy, N.K.; Mooers, A.O. Global priorities for conserving the evolutionary history of sharks, rays and chimaeras. Nat. Ecol. Evol. 2018, 2, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, B.; Kirkness, E.F.; Loh, Y.H.; Halpern, A.L.; Lee, A.P.; Johnson, J.; Dandona, N.; Viswanathan, L.D.; Tay, A.; Venter, J.C.; et al. Survey sequencing and comparative analysis of the elephant shark (Callorhinchus milii) genome. PLoS. Biol. 2007, 5, e101. [Google Scholar] [CrossRef] [PubMed]
- Aschliman, N.C.; Nishida, M.; Miya, M.; Inoue, J.G.; Rosana, K.M.; Naylor, G.J. Body plan convergence in the evolution of skates and rays (Chondrichthyes: Batoidea). Mol. Phylogenet. Evol. 2012, 63, 28–42. [Google Scholar] [CrossRef]
- Van Staden, M.; Ebert, D.A.; da Silva, C.; van der Bester Merwe, A.E. Comparative analyses of the complete mitochondrial genomes of two southern African endemic guitarfish, Acroteriobatus annulatus and A. blochii. Int. J. Biol. Macromol. 2022, 223, 1094–1106. [Google Scholar] [CrossRef]
- Chan, P.P.; Lin, B.Y.; Mak, A.J.; Lowe, T.M. tRNAscan-SE 2.0: Improved detection and functional classification of transfer RNA genes. Nucleic Acids Res. 2021, 49, 9077–9096. [Google Scholar] [CrossRef]
- Scharpf, C.; Lazara, K.J. The ETYFish project. Rhinopristiformes. Am. Curr. 2021, 1–6. [Google Scholar]
- Last, P.R.; De Carvalho, M.R.; Naylor, G.J.P.; Séret, B.; Stehmann, M.F.W. Rays of the World; CSIRO Publishing: Clayton South, VIC, Australia, 2016; Volume 790. [Google Scholar]
- Patterson, H.M.; D’Alberto, B.M.; Carlson, J.K.; Pardo, S.A.; Simpfendorfer, C.A. Population productivity of shovelnose rays: Inferring the potential for recovery. PLoS ONE 2019, 14, e0225183. [Google Scholar] [CrossRef]
- Marichamy, R. On a large-sized green saw fish Pristis zijsron Bleeker landed at Port Blair, Andamans. J. Mar. Biol. Assoc. India 1969, 10, 394. [Google Scholar]
- Weigmann, S. Annotated checklist of the living sharks, batoids and chimaeras (Chondrichthyes) of the world, with a focus on biogeographical diversity. J. Fish Biol. 2016, 88, 837–1037. [Google Scholar] [CrossRef]
- Wueringer, B.E. Sawfish captures in the Queensland Shark Control Program, 1962 to 2016. Endanger Species Res. 2017, 34, 293–300. [Google Scholar] [CrossRef]
- Lear, K.O.; Fazeldean, T.; Bateman, R.L.; Inglebrecht, J.; Morgan, D.L. Growth and morphology of Critically Endangered green sawfish Pristis zijsron in globally important nursery habitats. Mar. Biol. 2023, 170, 70. [Google Scholar] [CrossRef]
- Harry, A.V.; Everett, B.; Faria, V.; Fordham, S.; Grant, M.I.; Haque, A.B.; Ho, H.; Jabado, R.W.; Jones, G.C.A.; Lear, K.O.; et al. Pristis zijsron. The IUCN Red List of Threatened Species 2022: E.T39393A58304631. Mar. Biol. 2022, 170, 70. [Google Scholar] [CrossRef]
- Dulvy, N.K.; Davidson, L.N.K.; Kyne, P.M.; Simpfendorfer, C.A.; Harrison, L.R.; Carlson, J.K.; Fordham, S.V. Ghosts of the coast: Global extinction risk and conservation of sawfishes. Aquat. Conserv. Mar. Freshw. Ecosyst. 2014, 26, 134–153. [Google Scholar] [CrossRef]
- Leeney, R.H. Are sawfishes still present in Mozambique? A baseline ecological study. PeerJ. 2017, 5, e2950. [Google Scholar] [CrossRef]
- Simpfendorfer, C.A. Pristis zijsron (errata version published in 2019). In The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2013; E.T39393A141792003. [Google Scholar]
- Harrison, L.R.; Dulvy, N.K.E. Sawfish: A Global Strategy for Conservation; IUCN Species Survival Commission’s Shark Specialist Group: Vancouver, BC, Canada, 2014. [Google Scholar]
- Grant, M.I.; Charles, R.; Fordham, S.; Harry, A.V.; Lear, K.O.; Morgan, D.L.; Phillips, N.M.; Simeon, B.; Wakhida, Y.; Wueringer, B.E. Pristis clavata. In The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2022; e.T39390A68641215. [Google Scholar] [CrossRef]
- Carlson, J.; Blanco-Parra, M.P.; Bonfil-Sanders, R.; Charles, R.; Charvet, P.; Chevis, M.; Dulvy, N.K.; Espinoza, M.; Faria, V.; Ferretti, F.; et al. Pristis pectinata. In The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2022; E.T18175A58298676. [Google Scholar] [CrossRef]
- Espinoza, M.; Bonfil-Sanders, R.; Carlson, J.; Charvet, P.; Chevis, M.; Dulvy, N.K.; Everett, B.; Faria, V.; Ferretti, F.; Fordham, S.; et al. Pristis pristis. In The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2022; E.T18584848A58336780. [Google Scholar] [CrossRef]
- Henze, K.; Martin, W. Evolutionary biology: Essence of mitochondria. Nature 2003, 426, 127–128. [Google Scholar] [CrossRef]
- Muñoz-Gómez, S.A.; Wideman, J.G.; Roger, A.J.; Slamovits, C.H. The Origin of Mitochondrial Cristae from Alphaproteobacteria. Mol. Biol. Evol. 2017, 34, 943–956. [Google Scholar] [CrossRef]
- McBride, H.M.; Neuspiel, M.; Wasiak, S. Mitochondria: More than just a powerhouse. Curr. Biol. 2006, 16, R551–R560. [Google Scholar] [CrossRef]
- Wolstenholme, D.R. Animal Mitochondrial DNA: Structure and Evolution. Int. Rev. Cytol. 1992, 141, 173–216. [Google Scholar]
- Boore, J.L. Animal mitochondrial genomes. Nucleic. Acids. Res. 1999, 27, 1767–1780. [Google Scholar] [CrossRef]
- Steinke, D.; Bernard, A.M.; Horn, R.L.; Hilton, P.; Hanner, R.; Shivji, M.S. DNA analysis of traded shark fins and mobulid gill plates reveals a high proportion of species of conservation concern. Sci. Rep. 2017, 7, 9505. [Google Scholar] [CrossRef] [PubMed]
- Holmes, B.H.; Steinke, D.; Ward, R.D. Identification of shark and ray fins using DNA barcoding. Fish. Res. 2009, 95, 280–288. [Google Scholar] [CrossRef]
- Haque, A.B.; Das, S.A. First confirmed record of the Critically Endangered green sawfish Pristis zijsron from Bangladeshi waters. J. Fish. Biol. 2019, 94, 200–203. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.F. Molecular Cloning. A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989. [Google Scholar]
- Chen, X.; Kyne, P.M.; Pillans, R.D.; Feutry, P. Complete mitochondrial genome of the Endangered Narrow Sawfish Anoxypristis cuspidata (Rajiformes: Pristidae). Mitochondrial DNA A DNA Mapp. Seq. Anal. 2015, 27, 4172–4173. [Google Scholar] [CrossRef]
- Burland, T.G. DNASTAR’s Lasergene sequence analysis software. Methods. Mol. Biol. 2000, 132, 71–91. [Google Scholar]
- Bernt, M.; Donath, A.; Juhling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Putz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de novo metazoan mitochondrial genome annotation. Mol. Phylogenet. Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line: Integrating search and context for analysis of transfer RNA genes. Nucleic. Acids. Res. 2016, 44, 54–57. [Google Scholar] [CrossRef]
- Laslett, D.; Canback, B. ARWEN: A program to detect tRNA genes in metazoan mitochondrial nucleotide sequences. Bioinformatics 2008, 24, 172–175. [Google Scholar] [CrossRef]
- Wang, W. The Molecular Detection of Corynespora Cassiicola on Cucumber by PCR Assay Using DNAman Software and NCBI. In Computer and Computing Technologies in Agriculture IX; IFIP Advances in Information and Communication Technology; Springer: Berlin/Heidelberg, Germany, 2016; pp. 248–258. [Google Scholar]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic. Acids. Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef]
- Grant, J.R.; Stothard, P. The CGView Server: A comparative genomics tool for circular genomes. Nucleic. Acids. Res. 2008, 36, W181–W184. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sanchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Perna, N.T.; Kocher, T.D. Patterns of nucleotide composition at fourfold degenerate sites of animal mitochondrial genomes. J. Mol. Evol. 1995, 41, 353–358. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Ranwez, V.; Douzery, E.J.P.; Cambon, C.; Chantret, N.; Delsuc, F. MACSE v2: Toolkit for the Alignment of Coding Sequences Accounting for Frameshifts and Stop Codons. Mol. Biol. Evol. 2018, 35, 2582–2584. [Google Scholar] [CrossRef] [PubMed]
- Talavera, G.; Castresana, J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Johri, S.; Fellows, S.R.; Solanki, J.; Busch, A.; Livingston, I.; Mora, M.F.; Tiwari, A.; Cantu, V.A.; Goodman, A.; Morris, M.M.; et al. Mitochondrial genome to aid species delimitation and effective conservation of the Sharpnose Guitarfish (Glaucostegus granulatus). Meta Gene 2020, 24, 100648. [Google Scholar] [CrossRef]
- Feutry, P.; Kyne, P.M.; Grewe, P.M.; Chen, X.; Liu, M. Whole mitogenome of the Endangered dwarf sawfish Pristis clavata (Rajiformes: Pristidae). Mitochondrial DNA 2015, 26, 329–330. [Google Scholar] [CrossRef]
- Chen, X.; Wiley, T.; Kyne, P.M.; Feutry, P. Complete mitochondrial genome of the Critically Endangered Smalltooth Sawfish Pristis pectinata (Rajiformes: Pristidae). Mitochondrial DNA A DNA Mapp. Seq. Anal. 2015, 27, 3331–3332. [Google Scholar] [CrossRef]
- Kyne, P.M.; Wang, J.-J.; Xiang, D.; Chen, X.; Feutry, P. The phylogenomic position of the Critically Endangered Largetooth Sawfish Pristis pristis (Rhinopristiformes, Pristidae), inferred from the complete mitochondrial genome. Mitochondrial DNA Part B Resour. 2018, 3, 970–971. [Google Scholar] [CrossRef] [PubMed]
- Si, R.; Chen, H.; Ai, W.; Chen, X.; Chen, S. Complete mitochondrial genome and the phylogenetic position of the Bowmouth guitarfish Rhina ancylostoma (Rajiformes, Rhinobatidae). Mitochondrial DNA B Resour. 2016, 1, 270–272. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ai, W.; Shi, X.; Gao, T. Mitochondrial genome of the ringstraked guitarfish Rhinobatos hynnicephalus (Elasmobranchii: Rajiformes). Mitochondrial DNA 2015, 26, 653–654. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ai, W.; Xiang, D.; Pan, L.; Shi, X. Complete mitogenome of the brown guitarfish Rhinobatos schlegelii (Rajiformes, Rhinobatidae). Mitochondrial DNA A DNA Mapp. Seq. Anal. 2016, 27, 310–311. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Páez, A.; del Río-Portilla, M.A.; Oñate-González, E.; Rocha-Olivares, A. The mitochondrial genome of the banded guitarfish, Zapteryx exasperata (Jordan and Gilbert, 1880), possesses a non-coding duplication remnant region. Mitochondrial DNA 2014, 27, 1668–1670. [Google Scholar] [CrossRef]
- Si, R.; Ding, W.; Chen, H.; Chen, X.; Ai, W. Complete mitochondrial genome and the phylogenetic position of the White-spotted guitarfish Rhynchobatus australiae (Rajiformes, Rhinobatidae). Mitochondrial DNA B Resour. 2016, 1, 315–317. [Google Scholar] [CrossRef]
- Groeneveld, M.J.; Klein, J.D.; Bennett, R.H.; Bester-van der Merwe, A.E. Characterization of the complete mitochondrial genomes of two Critically Endangered wedgefishes: Rhynchobatus djiddensis and Rhynchobatus australiae. Mitochondrial DNA Part B Resour. 2023, 8, 352–358. [Google Scholar] [CrossRef]
- Johri, S.; Tiwari, A.; Kerr, E.N.; Dinsdale, E.A. Mitochondrial genome of the Smoothnose wedgefish Rhynchobatus laevis from the Western Indian Ocean. Mitochondrial DNA B Resour. 2020, 5, 2083–2084. [Google Scholar] [CrossRef]
- Inoue, J.G.; Miya, M.; Lam, K.; Tay, B.H.; Danks, J.A.; Bell, J.; Walker, T.I.; Venkatesh, B. Evolutionary origin and phylogeny of the modern holocephalans (Chondrichthyes: Chimaeriformes): A mitogenomic perspective. Mol. Biol. Evol. 2010, 27, 2576–2586. [Google Scholar] [CrossRef]
- Sati, J.; Goel, C.; Kumar, R.; Ali, S.; Patiyal, R.S.; Singh, V.K.; Sahoo, P.K.; Barat, A. Complete mitochondrial genome organization of Tor putitora. Mitochondrial DNA 2014, 25, 278–279. [Google Scholar] [CrossRef]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New Methods for Selecting Partitioned Models of Evolution for Molecular and Morphological Phylogenetic Analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods. 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchene, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kuhnert, D.; De Maio, N.; et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Suleski, M.; Hedges, S.B. TimeTree: A Resource for Timelines, Timetrees, and Divergence Times. Mol. Biol. Evol. 2017, 34, 1812–1819. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef]
- Wang, C.; Chen, H.; Tian, S.; Yang, C.; Chen, X. Novel Gene Rearrangement and the Complete Mitochondrial Genome of Cynoglossus monopus: Insights into the Envolution of the Family Cynoglossidae (Pleuronectiformes). Int. J. Mol. Sci. 2020, 21, 6895. [Google Scholar] [CrossRef]
- Last, P.R.; Kyne, P.M.; Compagno, L.J. A new species of wedgefish Rhynchobatus cooki (Rhinopristiformes, Rhinidae) from the Indo-West Pacific. Zootaxa 2016, 4139, 233–247. [Google Scholar] [CrossRef]
- Ebert, D.A.; Gon, O. Rhinobatos austini n. sp., a new species of guitarfish (Rhinopristiformes: Rhinobatidae) from the Southwestern Indian Ocean. Zootaxa 2017, 4276, 204–214. [Google Scholar] [CrossRef]
- Habib, K.A.; Islam, M.J. Description of a new species of giant guitarfish, Glaucostegus younholeei sp. nov. (Rhinopristiformes: Glaucostegidae) from the northern Bay of Bengal, Bangladesh. Zootaxa 2021, 4995, 129–146. [Google Scholar] [CrossRef] [PubMed]
- White, W.T.; Last, P.R.; Naylor, G.J. Rhinobatos manai sp. nov., a new species of guitarfish (Rhinopristiformes: Rhinobatidae) from New Ireland, Papua New Guinea. Zootaxa 2016, 4175, 588–600. [Google Scholar] [CrossRef] [PubMed]
- Last, P.R.; Seret, B.; Naylor, G.J.P. Description of Rhinobatos ranongensis sp. nov. (Rhinopristiformes: Rhinobatidae) from the Andaman Sea and Bay of Bengal with a review of its northern Indian Ocean congeners. Zootaxa 2019, 4576, zootaxa-4576. [Google Scholar] [CrossRef] [PubMed]
- Last, P.R.; Séret, B.; Naylor, G.J. A new species of guitarfish, Rhinobatos borneensis sp. nov. with a redefinition of the family-level classification in the order Rhinopristiformes (Chondrichthyes: Batoidea). Zootaxa 2016, 4117, 451–475. [Google Scholar] [CrossRef]
- Boaz, N.; El-Arnauti, A.; Gaziry, A.W.; Heinzelin, J.; Boaz, D. Neogene Paleontology and Geology of Sahabi, Libya; Alan R. Liss., Inc.: New York, NY, USA, 1987; pp. 145–151. [Google Scholar]
- Laurito, C.A. Los Selaceos Fósiles de Ia Localidad de Alto Guayacan (y Otros Ictiolitos Asociados); Texto Comunicación S.A.: San José, Costa Rica, 1999; p. 168. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Species | Length (bp) | AT% | GenBank ID | Reference |
|---|---|---|---|---|---|
| Glaucostegidae | Glaucostegus granulatus | 16,547 | 59.8 | MN783017 | [46] |
| Pristidae | Anoxypristis cuspidata | 17,243 | 61.3 | NC_026307 | [31] |
| Pristis clavata | 16,804 | 60.2 | NC_022821 | [47] | |
| Pristis pectinata | 16,802 | 61.0 | NC_027182 | [48] | |
| Pristis pristis | 16,912 | 60.3 | NC_039438 | [49] | |
| Pristis zijsron | 16,804 | 62.0 | MH005927 | This study | |
| Rhinidae | Rhina ancylostoma | 17,217 | 62.3 | NC_030215 | [50] |
| Rhinobatidae | Acroteriobatus annulatus | 16,773 | 59.8 | NC_068897 | [5] |
| Acroteriobatus blochii | 16,771 | 60.1 | NC_068898 | [5] | |
| Rhinobatos hynnicephalus | 16,776 | 59.8 | NC_022841 | [51] | |
| Rhinobatos schlegelii | 16,780 | 60.4 | NC_023951 | [52] | |
| Zapteryx exasperata | 17,310 | 62.5 | NC_024937 | [53] | |
| Rhynchobatidae | Rhynchobatus australiae | 16,804 | 59.8 | NC_030254 | [54] |
| Rhynchobatus djiddensis | 16,799 | 60.0 | NC_066688 | [55] | |
| Rhynchobatus laevis | 16,560 | 59.5 | NC_047241 | [56] | |
| Outgroup | Callorhinchus milii | 16,769 | 66.2 | NC_014285 | [57] |
| Tor putitora | 16,576 | 57.0 | NC_021755 | [58] |
| Region | Strand | Position | Size (bp) | Start Codon | Stop Codon | Intergenic Spacer |
|---|---|---|---|---|---|---|
| tRNA-Phe (F) | H | 1–69 | 69 | 0 | ||
| 12S rRNA | H | 70–1032 | 963 | 0 | ||
| tRNA-Val (V) | H | 1033–1105 | 73 | 0 | ||
| 16S rRNA | H | 1106–2799 | 1695 | 0 | ||
| tRNA-Leu (L) | H | 2800–2874 | 75 | 0 | ||
| ND1 | H | 2875–3849 | 975 | ATG | TAA | 2 |
| tRNA-Ile (I) | H | 3852–3921 | 70 | −1 | ||
| tRNA-Gln (Q) | L | 3921–3992 | 72 | 0 | ||
| tRNA-Met (M) | H | 3993–4062 | 70 | 0 | ||
| ND2 | H | 4063–5109 | 1047 | ATG | TAA | −1 |
| tRNA-Trp (W) | H | 5109–5177 | 69 | 2 | ||
| tRNA-Ala (A) | L | 5180–5248 | 69 | 0 | ||
| tRNA-Asn (N) | L | 5249–5321 | 73 | 0 | ||
| OL | - | 5322–5354 | 33 | 0 | ||
| tRNA-Cys (C) | L | 5355–5423 | 69 | 0 | ||
| tRNA-Tyr (Y) | L | 5424–5493 | 70 | 1 | ||
| COI | H | 5495–7051 | 1557 | GTG | TAA | 4 |
| tRNA-Ser (S) | L | 7056–7126 | 71 | 1 | ||
| tRNA-Asp (D) | H | 7128–7195 | 68 | 6 | ||
| COII | H | 7202–7892 | 691 | ATG | T- | 0 |
| tRNA-Lys (K) | H | 7893–7966 | 74 | 1 | ||
| ATP8 | H | 7968–8135 | 168 | ATG | TAA | −10 |
| ATP6 | H | 8126–8809 | 684 | ATG | TAA | 6 |
| COIII | H | 8816–9601 | 786 | ATG | TAA | 1 |
| tRNA-Gly (G) | H | 9603–9672 | 70 | 0 | ||
| ND3 | H | 9673–10,021 | 349 | ATG | T- | 1 |
| tRNA-Arg (R) | H | 10,023–10,092 | 70 | 1 | ||
| ND4L | H | 10,094–10,390 | 297 | ATG | TAA | −7 |
| ND4 | H | 10,384–11,764 | 1381 | ATG | T- | 0 |
| tRNA-His (H) | H | 11,765–11,833 | 69 | 0 | ||
| tRNA-Ser (S) | H | 11,834–11,900 | 67 | −1 | ||
| tRNA-Leu (L) | H | 11,900–11,971 | 72 | 0 | ||
| ND5 | H | 11,972–13,810 | 1839 | ATG | TAA | −4 |
| ND6 | L | 13,807–14,325 | 519 | ATG | TAG | 1 |
| tRNA-Glu (E) | L | 14,327–14,395 | 69 | 3 | ||
| Cytb | H | 14,399–15,541 | 1143 | ATG | TAA | 4 |
| tRNA-Thr (T) | H | 15,546–15,619 | 74 | 5 | ||
| tRNA-Pro (P) | L | 15,625–15,694 | 70 | 0 | ||
| Control region | - | 15,695–16,804 | 1110 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Ye, P.; Pillans, R.; Chen, X.; Wang, J.; Feutry, P. Evolution of the Critically Endangered Green Sawfish Pristis zijsron (Rhinopristiformes, Pristidae), Inferred from the Whole Mitochondrial Genome. Genes 2023, 14, 2052. https://doi.org/10.3390/genes14112052
Wang C, Ye P, Pillans R, Chen X, Wang J, Feutry P. Evolution of the Critically Endangered Green Sawfish Pristis zijsron (Rhinopristiformes, Pristidae), Inferred from the Whole Mitochondrial Genome. Genes. 2023; 14(11):2052. https://doi.org/10.3390/genes14112052
Chicago/Turabian StyleWang, Chen, Peiyuan Ye, Richard Pillans, Xiao Chen, Junjie Wang, and Pierre Feutry. 2023. "Evolution of the Critically Endangered Green Sawfish Pristis zijsron (Rhinopristiformes, Pristidae), Inferred from the Whole Mitochondrial Genome" Genes 14, no. 11: 2052. https://doi.org/10.3390/genes14112052