
Clinical and Pathological Findings Associated with Mycobacteriosis in Captive Syngnathids
 , , ,
, , ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Case Selection
2.2. Animal Husbandry Conditions
2.3. Necropsy and Histology
2.4. Molecular Identification
3. Results
3.1. Clinical Signs and Management
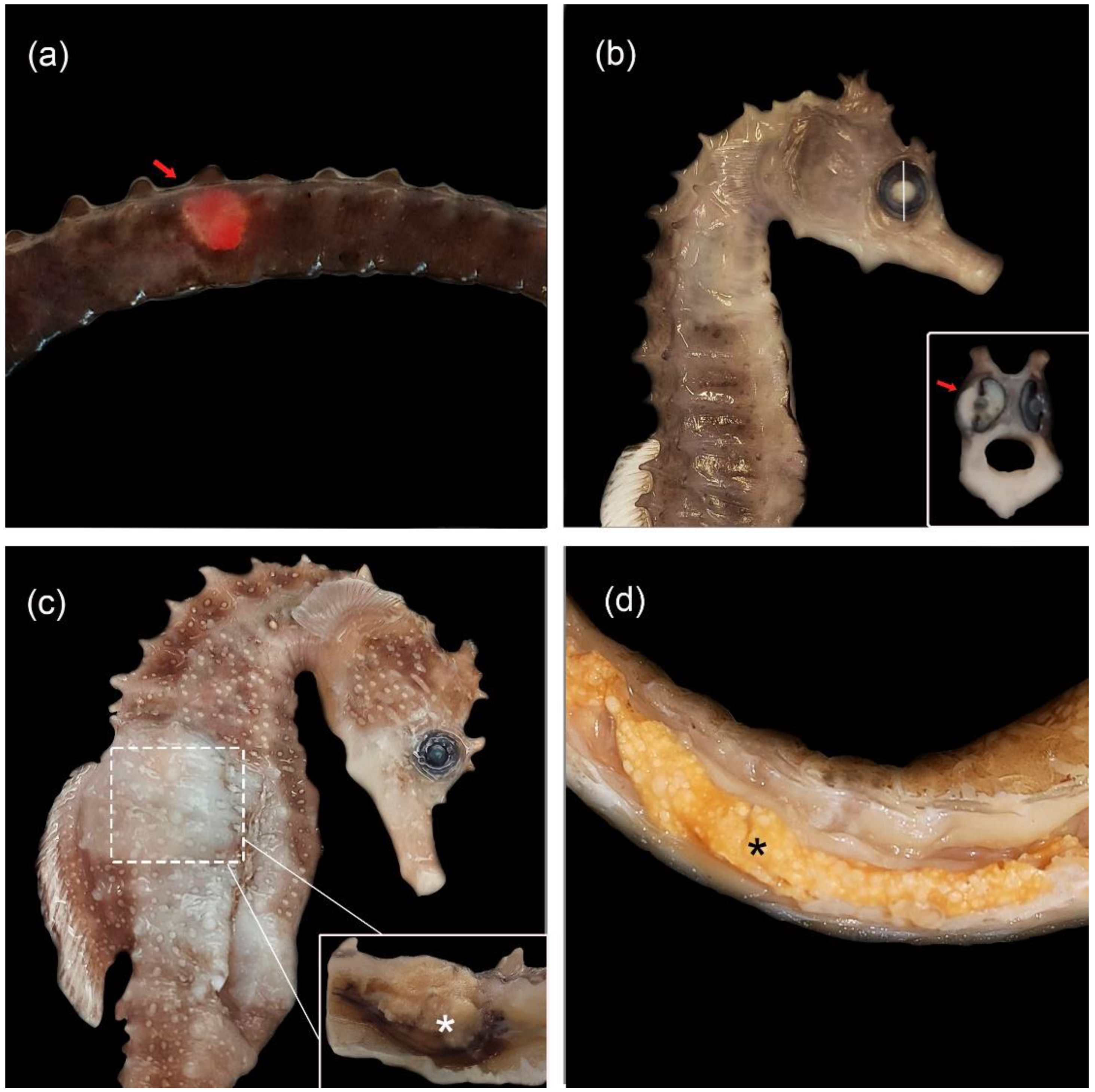
3.2. Gross and Histopathological Findings
3.3. Bacterial Identification by PCR
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jacobs, J.M.; Stine, C.B.; Baya, A.M.; Kent, M.L. Review of mycobacteriosis in marine fish. J. Fish Dis. 2009, 32, 119–130. [Google Scholar] [CrossRef]
- Reza, M.; El-Matbouli, M.; Menanteau-Ledouble, S. Mycobacteriosis and Infections with Non-tuberculous Mycobacteria in Aquatic Organisms: A Review. Microorganisms 2020, 8, 1368. [Google Scholar]
- Fogelson, S.; Fast, M.D.; Leary, J.; Camus, A.C. Pathologic features of mycobacteriosis in naturally infected Syngnathidae and novel transcriptome assembly in association with disease. J. Fish Dis. 2017, 40, 1681–1694. [Google Scholar] [CrossRef]
- Gauthier, D.T.; Rhodes, M.W. Mycobacteriosis in fishes: A review. Vet J. 2009, 180, 33–47. [Google Scholar] [CrossRef]
- Puk, K.; Banach, T.; Wawrzyniak, A.; Adaszek, Ł.; Zietek, J.; Winiarczyk, S.; Guz, L. Detection of Mycobacterium marinum, M. peregrinum, M. fortuitum and M. abscessus in aquarium fish. J. Fish Dis. 2018, 41, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Beran, V.; Matlova, L.; Dvorska, L.; Svastova, P.; Pavlik, I. Distribution of mycobacteria in clinically healthy ornamental fish and their aquarium environment. J. Fish Dis. 2006, 29, 383–393. [Google Scholar] [CrossRef]
- Balcázar, J.L.; Planas, M.; Pintado, J. Novel mycobacterium species in seahorses with tail rot. Emerg. Infect. Dis. 2011, 17, 1770–1772. [Google Scholar] [CrossRef] [Green Version]
- Mugetti, D.; Tomasoni, M.; Pastorino, P.; Esposito, G.; Menconi, V.; Dondo, A.; Prearo, M. Gene Sequencing and Phylogenetic Analysis: Powerful Tools for an Improved Diagnosis of Fish Mycobacteriosis Caused by Mycobacterium fortuitum Group Members. Microorganisms 2021, 9, 797. [Google Scholar] [CrossRef]
- Ortega, J.; Noguera, A.; García-Quirós, A.; Viana, D.; Selva, L.; de Juan, L.; Romero, B.; García-Parraga, D.; Crespo, J.L.; Corpa, J.M. Lesional patterns associated with mycobacteriosis in an Atlantic horse mackerel, Trachurus trachurus (L.), aquarium population. J. Fish Dis. 2014, 37, 591–595. [Google Scholar] [CrossRef] [PubMed]
- Stockton, J.; Torres, A. Multinucleated Giant Cell Formation as a Portal to Chronic Bacterial Infections. Microorganisms 2020, 8, 1637. [Google Scholar] [CrossRef]
- LePage, V.; Young, J.; Dutton, C.J.; Crawshaw, G.; Pare, J.A.; Kummrow, M.; McLelland, D.J.; Huber, P.; Young, K.; Russell, S.; et al. Diseases of captive yellow seahorse Hippocampus kuda Bleeker, pot-bellied seahorse Hippocampus abdominalis Lesson and weedy seadragon Phyllopteryx taeniolatus (Lacépède). J. Fish Dis. 2014, 38, 439–450. [Google Scholar] [CrossRef]
- Bonar, C.J.; Garner, M.M.; Weber, E.S.; Keller, C.J.; Murray, M.; Adams, L.M.; Frasca, S. Pathologic Findings in Weedy (Phyllopteryx taeniolatus) and Leafy (Phycodurus eques) Seadragons. Vet. Pathol. 2013, 50, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Foster, S.; Wiswedel, S.; Vincent, A. Opportunities and challenges for analysis of wildlife trade using CITES data-seahorses as a case study. Aquat. Conserv. 2016, 26, 154–172. [Google Scholar] [CrossRef]
- Randazzo, B.; Rolla, L.; Ofelio, C.; Planas, M.; Gioacchini, G.; Vargas, A.; Giorgini, E.; Olivotto, I. The influence of diet on the early development of two seahorse species (H. guttulatus and H. reidi): Traditional and innovative approaches. Aquaculture 2018, 490, 75–90. [Google Scholar] [CrossRef]
- Foster, S.J.; Vincent, A.C.J. Life history and ecology of seahorses: Implications for conservation and management. J. Fish Biol. 2004, 65, 1–61. [Google Scholar] [CrossRef]
- Rosa, I.L.; Oliveira, T.P.R.; Osório, F.M.; Moraes, L.; Catro, A.; Barros, G.; Mulo, R.; Alves, R. Fisheries, and trade of seahorses in Brazil: Historical perspective, current trends, and future directions. Biodivers. Conserv. 2011, 20, 1951–1971. [Google Scholar] [CrossRef]
- International Union for Conservation of Nature (IUCN). IUCN Red List of Threatened Species. IUCN. Available online: www.redlist.org (accessed on 23 May 2022).
- Koldewey, H.J.; Martin-Smith, K.M. A global review of seahorse aquaculture. Aquaculture 2010, 302, 131–152. [Google Scholar] [CrossRef]
- Prosser, C.M.; Unger, M.A.; Vogelbein, W.K. Multistressor interactions in the zebrafish (Danio rerio): Concurrent phenanthrene exposure and Mycobacterium marinum infection. Aquat. Toxicol. 2011, 102, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Fogelson, S.; Camus, A.; Lorenz, W.; Phillips, A.; Barlett, P.; Sanchez, S. Mycobacterium syngnathidarum sp. nov., a rapidly growing mycobacterium identified in syngnathid fish. Int. J. Syst. Evol. Microbiol. 2018, 68, 3696–3700. [Google Scholar] [CrossRef] [PubMed]
- Garrido, J.M.; Cortabarria, N.; Oguiza, J.A.; Aduriz, G.; Juste, R.A. Use of a PCR method on fecal samples for diagnosis of sheep paratuberculosis. Vet. Microbiol. 2000, 77, 379–386. [Google Scholar] [CrossRef]
- Bartos, M.; Hlozek, P.; Svastova, P.; Dvorska, L.; Bull, T.; Parmova, I.; Kuhn, I.; Stubbs, J.; Moravkova, M.; Kintr, J.; et al. Identification of members of Mycobacterium avium species by Accu-Probes, serotyping, and single IS900, IS901, IS1245 and IS901-flanking region PCR with internal standards. J. Microbiol. Methods 2006, 64, 333–345. [Google Scholar] [CrossRef] [PubMed]
- Parikka, M.; Hammarén, M.; Harjula, S.-K.; Halfpenny, N.; Oksanen, K.; Lahtinen, M.; Pajula, E.; Iivanainen, A.; Pesu, M.; Rämet, M. Mycobacterium marinum causes a latent infection that can be reactivated by gamma irradiation in adult zebrafish. PLoS Pathog. 2012, 8, e1002944. [Google Scholar] [CrossRef] [Green Version]
- Phung, T.N.; Caruso, D.; Godreuil, S.; Keck, N.; Vallaeys, T.; Avarre, C. Rapid detection and identification of nontuberculous mycobacterial pathogens in fish by using high-resolution melting analysis. Appl. Environ. Microbiol. 2013, 79, 7837–7845. [Google Scholar] [CrossRef] [Green Version]
- Hughes, M.S.; James, G.; Taylor, M.J.; McCarroll, J.S.; Neill, D.; Chen, S.C.A.; Mitchell, D.H.; Love, N.; Malik, R. PCR studies of feline leprosy cases. J. Feline Med. Surg. 2004, 6, 235–243. [Google Scholar] [CrossRef]
- Esfahani, B.N.; Yazdi, H.; Moghim, S.; Safaei, H.; Zarkesh, H.; Esfahani, Z. Rapid and accurate identification of Mycobacterium tuberculosis complex and common non-tuberculous mycobacteria by multiplex real-time PCR targeting different housekeeping genes. Curr. Microbiol. 2012, 65, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Novotny, L.; Halouzka, R.; Matlova, L.; Vavra, O.; Bartosova, L.; Slany, M.; Pavlik, I. Morphology and distribution of granulomatous inflammation in freshwater ornamental fish infected with mycobacteria. J. Fish Dis. 2010, 33, 947–955. [Google Scholar] [CrossRef]
- Kim, S.H.; Shin, J.H. Identification of nontuberculous mycobacteria using multilocous sequence analysis of 16S rRNA, hsp65, and rpoB. J. Clin. Lab. Anal. 2018, 32, e22184. [Google Scholar] [CrossRef] [Green Version]
- Morais, F.; Bello, G.M.; Costi, C.; Schmid, K.B.; Dos Santos-Soares, T.; Bones, R.; Unis, G.; Fontoura, C.; Almeida, P.E.; Rossetti, M.L. Detection of non-tuberculosus mycobacteria (NTMs) in lung samples using 16S rRNA. Mem. Inst. Oswaldo Cruz 2022, 117, e220031. [Google Scholar] [CrossRef]
- Yanong, R.P.; Curtis, E.W.; Terrell, S.P.; Case, G. Atypical presentation of mycobacteriosis in a collection of frogfish (Antennarius striatus). J. Zoo Wildl. Med. 2003, 34, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Gcebe, N.; Michel, A.L.; Hlokwe, T.M. Non-tuberculous Mycobacterium species causing mycobacteriosis in farmed aquatic animals of South Africa. BMC Microbiol. 2018, 18, 32. [Google Scholar] [CrossRef]
- Loeschke, S.; Goldmann, T.; Vollmer, E. Improved detection of mycobacterial DNA by PCR in formalin-fixed, paraffin-embedded tissues using thin sections. Pathol. Res. Pract. 2005, 201, 37–40. [Google Scholar] [CrossRef] [PubMed]
- Barcelos, D.; Franco, M.F.; Leão, S.C. Effects of tissue handling and processing steps on PCR for detection of Mycobacterium tuberculosis in formalin-fixed paraffin-embedded samples. Rev. Inst. Med. Trop. Sao Paulo 2008, 50, 321–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavlik, I.; Ulmann, V.; Falkinham, J.O. Nontuberculous Mycobacteria: Ecology and Impact on Animal and Human Health. Microorganisms 2022, 10, 1516. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Species (Gene) | Primer Design |
|---|---|
| Mycobacterium avium subsp. paratuberculosis (IS900) [21] | F:GATCGGAACGTCGGCTGGTCAGG |
| R:GATCGCCTTGCTCATCGCTGCCG | |
| Mycobacterium avium subsp. avium (IS901) [22] | F:AAGCCGAGGTGGTGTATGT |
| R:AGCGAAGATGGCGGTGAGCAT | |
| Mycobacterium marinum (16S-23S ITS) [23] | F:CACCACGAGAAACACTCCAA |
| R:ACATCCCGAAACCAACAGAG | |
| M marinum-M fortuitum-M chelonae (16S-23S ITS) [24] | F:GCTGGATCACCTCCTTTCTA |
| R:AGATGCTCGCAACCACTAT | |
| M lepraemurium (16S rRNA) [25] | F:GAATATTGCACAATGGGCGCAG R: AAACCCGGACCTTCGTCGATA |
| oneM fortuitum (16S-23S ITS) [26] | F:GACTGCCAGACACACTATTGG |
| R:GTGAGACCACACGATTCTGC |
| Syngnathids | Species | Sex | Date of Sampling | Acute or Chronic Presentation | Clinical Signs and Gross Changes | Affected Tissue | Bacterial Load | Identified Microorganism | |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Seahorse | Hippocampus guttulatus | Female | 03/11/2020 | Chronic | Dyspnea mucus in gills, tail ulcer | Liver, skin, kidney, gills, intestine, heart | Mild to moderate | M chelonae, M marinum |
| 2 | Hippocampus guttulatus | Female | 30/09/2020 | Acute | Sudden death | Liver, intestine, heart | Moderate | M fortuitum | |
| 3 | Hippocampus guttulatus | Male | 19/02/2021 | Chronic | Dyspnea, mucus in gills | Liver, heart, intestine, gills | Moderate | M chenolae, M marinum | |
| 4 | Hippocampus guttulatus | Male | 17/02/2021 | Chronic | Dyspnea, mucus in gills tail ulcer, poor body condition | Liver, intestine, skeletal muscle, kidney, skin, gills, heart, blood vessel | Moderate | M chelonae, M marinum | |
| 5 | Hippocampus guttulatus | Female | 17/03/2021 | Chronic | Dyspnea, mucus in gills, white eye, exophthalmia | Liver, intestine, kidney, gills, eye | Mild to moderate | M fortuitum | |
| 6 | Hippocampus guttulatus | Male | 24/05/2021 | Acute | Sudden death | Liver, intestine, gills, heart | Moderate | M chenolae, M marinum | |
| 7 | Hippocampus guttulatus | Male | 05/09/2021 | Chronic | Tail ulcer, soft exoskeleton, mucus in gills | Liver, intestine, gills, heart, skin | Mild to moderate | Mycobacterium sp. | |
| 8 | Hippocampus guttulatus | Male | 11/08/2021 | Chronic | Coelomic cavity mass, poor body condition | Liver, intestine, skeletal muscle, kidney, gills | Mild to moderate | M chenolae, M marinum | |
| 9 | Hippocampus guttulatus | Male | 25/08/2021 | Chronic | White eye, exophthalmia, mucus in gills | Liver, intestine, skeletal muscle, kidney, skin, heart, gills, blood vessel, brain | Moderate | M chenolae, M marinum | |
| 10 | Hippocampus reidi | Female | 08/12/2020 | Acute | Sudden death | Gills, blood vessel | Moderate to severe | Mycobacterium sp. | |
| 11 | Hippocampus reidi | Female | 13/01/2021 | Acute | Sudden death | Liver, gills | Moderate | Mycobacterium sp. | |
| 12 | Hippocampus reidi | Male | 02/02/2021 | Chronic | Poor body condition | Liver, intestine, gills, blood vessel | Moderate | Mycobacterium sp. | |
| 13 | Hippocampus reidi | Male | 14/4/2021 | Chronic | Soft exoskeleton | Liver, intestine, skeletal muscle, gills, brain | Moderate | Mycobacterium sp. | |
| 14 | Hippocampus reidi | Female | 12/03/2021 | Chronic | Poor body condition | Liver, intestine, kidney, gills, brain | Mild to moderate | Mycobacterium sp. | |
| 15 | Hippocampus reidi | Female | 18/08/2021 | Chronic | Tail ulcer, soft exoskeleton, mucus in gills | Kidney, skin, gills, brain, blood vessel | Moderate | Mycobacterium sp. | |
| 16 | Hippocampus reidi | Female | 16/09/2021 | Chronic | Dyspnea, mucus in gills | Liver, kidney, gills, brain | Moderate | Mycobacterium sp. | |
| 17 | Hippocampus abdominalis | Male | 18/05/2018 | Acute | Sudden death | Liver, intestine, gills | Moderate | Mycobacterium sp. | |
| 18 | Hippocampus abdominalis | Female | 11/07/2018 | Chronic | Poor body condition | Gills, eye, swim bladder | Mild to moderate | Mycobacterium sp. | |
| 19 | Hippocampus abdominalis | Female | 26/02/2021 | Acute | Sudden death | Gills, blood vessel | Moderate to severe | Mycobacterium sp. | |
| 20 | Hippocampus abdominalis | Male | 23/04/2021 | Acute | Sudden death | Gills, blood vessel | Moderate | M fortuitum | |
| 21 | Pipe fish | Syngnathus acus | Female | 11/07/2018 | Acute | Sudden death | Gills, blood vessel | Moderate | Mycobacterium sp. |
| 22 | Syngnathus acus | Female | 05/04/2021 | Chronic | Soft exoskeleton | Liver, intestine, gills, gas gland | Mild to moderate | Mycobacterium sp. | |
| 23 | Syngnathoides biaculeatus | Male | 16/07/2018 | Acute | Sudden death | Gills, blood vessel | Moderate | M chenolae, M marinum | |
| 24 | Seadragon | Phyllopteryx taeniolatus | Female | 8/07/2018 | Chronic | Poor body condition | Liver, kidney, ovary | Mild to moderate | Mycobacterium sp. |
| 25 | Phyllopteryx taeniolatus | Female | 25/01/2021 | Chronic | Poor body condition | Liver, intestine, gills, heart, kidney | Mild to moderate | M fortuitum | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montero, E.; Rojo-Solís, C.; de Castro, N.; Fernández, M.; Pérez, V.; Corpa, J.M.; Ortega, J. Clinical and Pathological Findings Associated with Mycobacteriosis in Captive Syngnathids. Animals 2022, 12, 3259. https://doi.org/10.3390/ani12233259
Montero E, Rojo-Solís C, de Castro N, Fernández M, Pérez V, Corpa JM, Ortega J. Clinical and Pathological Findings Associated with Mycobacteriosis in Captive Syngnathids. Animals. 2022; 12(23):3259. https://doi.org/10.3390/ani12233259
Chicago/Turabian StyleMontero, Estefanía, Carlos Rojo-Solís, Noelia de Castro, Miguel Fernández, Valentín Pérez, Juan M. Corpa, and Joaquín Ortega. 2022. "Clinical and Pathological Findings Associated with Mycobacteriosis in Captive Syngnathids" Animals 12, no. 23: 3259. https://doi.org/10.3390/ani12233259